
INTRODUCTION
Plasmodium falciparum merozoites invade red blood cells (RBC), and over 48 hours develop and multiply through the ring, trophozoite and schizont stages [1, 2]. This asexual erythrocytic stage of the life cycle is attractive to study because it is responsible for the symptoms of the disease, and can be cultivated in vitro. The genome sequence has contributed enormously to our understanding of molecular and cellular aspects of the life cycle [3], however gene regulation in P. falciparum is still poorly understood.
Few transcription factors or regulatory motifs have been identified and characterized in malaria parasites [4-6]. Gene expression analysis revealed that transcription is generally monocistronic and finely regulated [7-12]. The AT-rich P. falciparum genome and the complex life cycle involving differential gene expression at morphologically distinct stages make it difficult to predict and to characterize transcription factors. Analysing the genome revealed a relative paucity of transcription-associated proteins and specific cis-regulator motifs [13], but the regulatory machinery includes a canonical TATA-box binding protein and RNA polymerase II-dependent messenger RNA production [14, 15].
Using bioinformatics, including genomic and proteomic tools, some orthologues of general transcription factors such as the TFIIB family [15] have been predicted. Genes encoding transcriptional regulators such as the Myb1 protein [16], high mobility group box (HMGB) proteins, and Apetala2 (AP2) [17] have been characterized in P. falciparum, and recently a new set of 129 cis-regulatory elements was reported [18].
Our group has studied for more than a decade how melatonin, a tryptophan-derived metabolite, modulates the cycle of P. falciparum and P. chabaudi parasites [19, 20]. A signaling cascade is induced by the indolamine, thus prompting a rise in the second messengers IP3, Ca2+ and cyclic AMP [21-23]. cAMP activates protein kinase A (PKA), and consequently modulates anion conductance and vesicle trafficking in P. falciparum [24-26]. A variety of regulatory factors are modulated by second messenger signaling in vertebrates [27-30]. It is a feature of cell biology that Ca2+ has a role regulating long-term cell adaptation by controlling gene expression (for review see [31]).
On PlasmoDB database (http://www.plasmodb.org), Pf11_0477 (now described as PF3D7_1146600) transcript was predicted to have around 3906 bp according to northern blotting results [32]. Moreover, in the rodent malaria parasites P. berghei, has been identified a Pf Nf-YB orthologue named ORP1 (Oocyst Rupture Protein 1) also containing an histone-fold domain (HFD) NF_YB-like. The authors found that mutant parasites in which orp1 gene has been deleted, oocyst rupture, normally occurring in mosquito midgut around day 12 post blood meal, is inhibited but growth of asexual stages was not affected [33]. The rupture of the oocyst is an essential step for parasite transmission and ORP1 plays a fundamental role in this mechanism.
We have demonstrated that melatonin induces posttranslational modification of the PfNF-YB protein by increasing its ubiquitination, allowing PfNF-YB abundance to be controlled by the ubiquitin-proteasome system [34]. In transcriptional studies PfNF-YB was detected in asexual schizont and in sexual gametocyte stages [14, 35]. The NF-Y protein complex binds a CCAAT-box and in P. falciparum is comprised of three subunits A (Pf13_0043), B and C (Pf14_0374) [14]. In other organisms the NF-Y subunit B recognises and binds to the CCAAT sequence in the promoter region of various genes [36-38]. For example, NF-YB binds to mammalian genes during the G2/M phase of the cell cycle and regulates the expression of topoisomerase IIα, cyclin B1, CDC25C, E2F, CDC2 and thymidine kinase [39-41]. In contrast, in P. falciparum, the target gene(s) bound by PfNF-YB during the parasite cycle life have not been identified, and the role of this transcription factor is unknown. Our group reported that a cAMP analogue (6-Bnz-cAMP) modulates PfNF-YB transcript and proteins levels, raising the possibility of a role for this protein in parasite signaling pathways [34].
In this research, we identified several DNA binding sites candidates for PfNF-YB transcription factor at the schizont stage. Our results suggest that PfNF-YB is involved in the control of numerous different biological functions in the parasite, including protein translation folding and modification, intracellular transport, nucleic acid metabolism and cell redox homeostasis.
RESULTS
PfNF-YB is expressed at higher levels in mature forms of the intraerythrocytic parasite
In order to investigate PfNF-YB function during the P. falciparum asexual stage we followed its transcript expression throughout the intraerythrocytic developmental cycle using a synchronized parasite population. Figure 1 shows the RT-PCR results after analysing the transcripts at 5 different time-points of development up to 48-hours post invasion (hpi). The analysis revealed that PfNF-YB transcript abundance peaks at 44 hpi followed by a drastic drop in abundance at 48 hpi. Following invasion, the PfNF-YB transcript level is low at early ring stage (10 hpi) and slowly increases during parasite maturation. Our quantitative PCR results are similar in pattern to those in the microarray database available at http://plasmodb.org and published for P. falciparum 3D7 strain [42]. The higher expression of PfNF-YB in mature parasites led us to postulate that the protein might regulate genes involved in DNA replication and cell division.
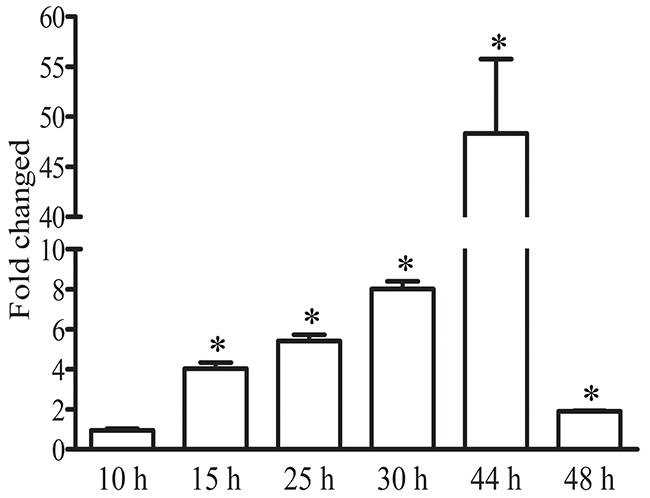
Figure 1: PfNF-YB expression throughout the intraerythrocytic cycle of P. falciparum. Samples of a synchronized parasite population were collected at various time points (10-48 h) after invasion. RT-PCR analysis of PfNF-YB mRNA level reveals differential expression during the intraerythrocytic cycle. Statistical analysis was performed by t test (p<0.05). The amount of mRNA in each sample was normalized by the level of 18S RNA.
The PfNF-YB transcription factor binds to P. falciparum DNA sequences
In various different eukaryotic cells NF-Y proteins are involved in cell cycle control, regulating several genes [40, 41]. We have shown here that P. falciparum NF-YB is highly expressed in late stages and located at least in part in the nucleus of both trophozoite and schizont forms. During these developmental stages, DNA duplication, followed by nuclear division and cytokinesis to produce new merozoites is essential to allow successful parasite multiplication and invasion of new red blood cells. Therefore, we asked whether PfNF-YB at the schizont stage (44 hpi) can bind to DNA and regulate expression of target genes involved in the parasite cell cycle. To answer this question, a genome-wide PfNF-YB chromatin immunoprecipitation (ChIP-on-chip) assay was performed using a high density whole genome tiling array (Roche NimbleGen Inc) and material purified from schizonts. Chromatin immunoprecipitation was done using PfNF-YB-specific rabbit antibody attached to beads, and, as controls, non-specific rabbit IgG attached to beads and beads alone. Before performing the ChiP-on-chip assay, qPCR with selected genes (PFI1665w and Pf14_0489) was used to test whether an enrichment of promoter regions was obtained using the PfNF-YB antibody. Using this approach, we observed that the putative promoter region of both PFI1665w and Pf14_0489 was enriched in the fraction bound to the PfNF-YB-specific antibody compare to the fraction bound to nonspecific rabbit IgG or beads alone (Supplementary Figure 1). For the more extensive ChiP-on-chip experiment, to determine the target genes for PfNF-YB binding we selected target sequences using three criteria: 1) the minimum Log 2 ratio value was set at 0.5; 2) the maximum false discovery rate (FDR) value was set at 0.095 (0.095 % FDR cut-off); and 3) probes covering more than 150 bp sequence were required (at least 4 probes, each probe of 50 bp). In the analysis we took into account the potential presence of putative promoter regions in the first exon and in other intragenic regions, since some genes in P. falciparum such as the var gene family have a promoter within the gene [43-45].
The graphic display generated by the Deva software shows PfNF-YB binding sites distributed in several putative promoter regions of target genes (Figure 2) throughout the 14 chromosomes of the P. falciparum genome (Figure 2A). PfNF-YB binding was homogeneously distributed on all chromosomes. A higher resolution display of ChiP-on-chip results for PfNF-YB binding to Chromosome7 is shown in Figure 2B. The candidate PfNF-YB target genes were selected by an enrichment above Log2 ≥ 0.5, a threshold that is represented in the figure by a dotted red line (Figure 2A and 2B). We exemplify these results using the MAL7P1.25 and PFB0325c target genes in Supplementary Figure 2. The Deva program established the peaks by searching for 4 or more probes whose signals were above the specified cutoff values, ranging from 90% to 15%, using a 500 bp sliding window. The cutoff values were a percentage of a hypothetical maximum, which was the mean + 6 standard deviations. The ratio data were then randomized 20 times to evaluate the probability of “false positives”. Each peak was then assigned a false discovery rate (FDR) score based on the randomization. We observed that the positive peaks for MAL7P1.25 and PFB0325c (Supplementary Figure 2) do not possess a high log2 ratio value and this may be due to the high AT content of the P. falciparum genome. AT-rich prokaryotic genomes may have less precise control of transcription initiation [46, 47], a phenomenon which may explain the low signal to noise ratio in our ChIP-on-chiP data. Inevitably this background will decrease the ease of definition of specific peaks, but it does not preclude the specificity of antibody binding to the transcription factor and of the transcription factor binding to target genes. As shown in Supplementary Table 1 we noted 297 target genes bound by PfNF-YB that fulfilled all three selection criteria.
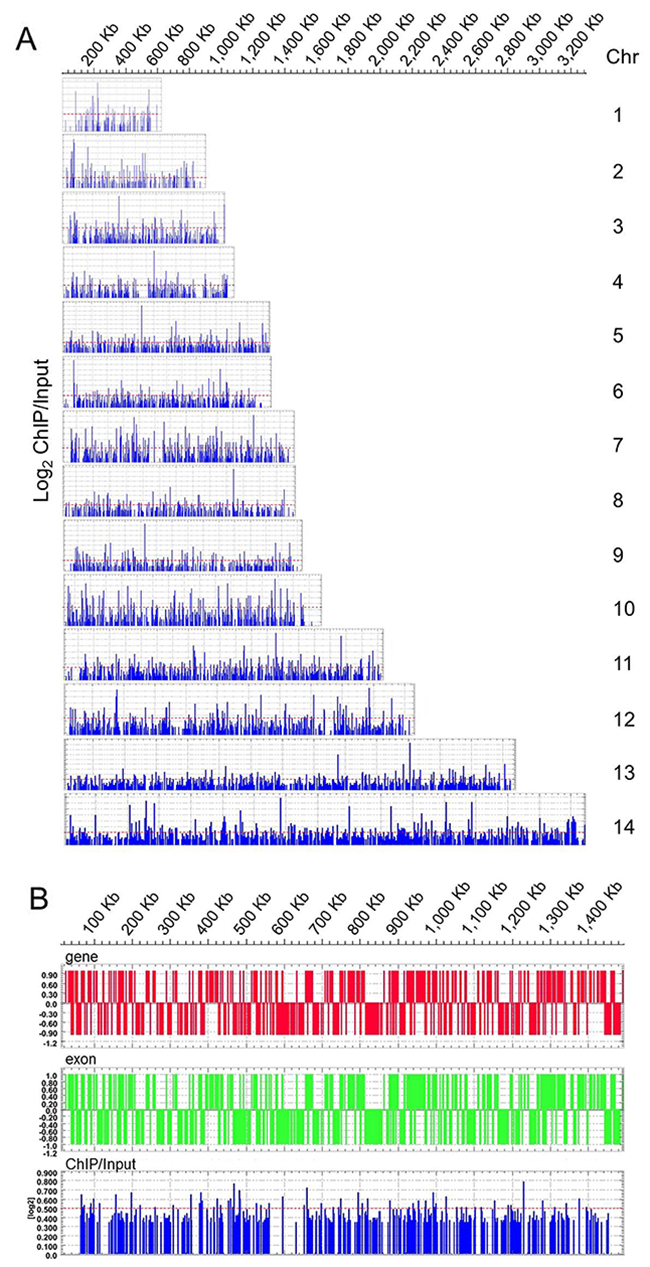
Figure 2: Genome-wide PfNF-YB occupancy by high-density ChIP-on-chip assay. The display (Deva) shows the chromosomal map position and location of DNA enrichment due to PfNF-YB binding. (A) The blue panel peaks shows the ChiP/Input ratio for all 14 chromosome in schizont stage 3D7 parasites. PfNF-YB occupancy for each gene was calculated as the average of log2 ratios of hybridization values for immunoprecipitated and input chromatin. (B). PfNF-YB occupancy on chromosome 7. Schematic display (Deva) of the blue peaks on chromosome 7 shows PfNF-YB occupancy in the gene region (red bars) and the exon region (green bars).The candidate promoter regions were estimated from false discovery rate (FDR) values (0.095) and by Log2 value ratio ≥ 0.5 (dotted red line). The ratio data were randomized 20 times to evaluate the probability of “false positives”. Each peak was assigned an FDR score based on the randomization. Chromosome numbers are indicated on the right, chromosomal position (Kb) on top.
Validation of target genes
Signal enrichment and FDR values are criteria that suggest binding regions, but in order to properly assess the array results, it was crucial to validate the targets identified from the ChIP-on-chip assay. Therefore, we used 3 different ChIP assays from schizonts (44 hpi) to confirm the enrichment and to measure the sensitivity and specificity of the detection by qPCR (Figure 3). Potential PfNF-YB targets were selected according to their enrichment, FDR value and biological function. We also chose three different regions with poor enrichment as controls in the qPCR using ChIP DNA. The analysis demonstrated that the selected 21 genes were positively enriched as judged by qPCR following immunoprecipitation with the PfNF-YB-specific antibody beads compare with the control immunoprecipitations. The three control genes MAL13P1.495, PFD0740w and MAL6P1.1 that were not enriched did not show any difference by qPCR between the PfNF-YB antibody and control bead immunoprecipitations. The qPCR method was shown to be more sensitive to detect enrichment than the ChIP-on-chip approach, once candidates had been identified.
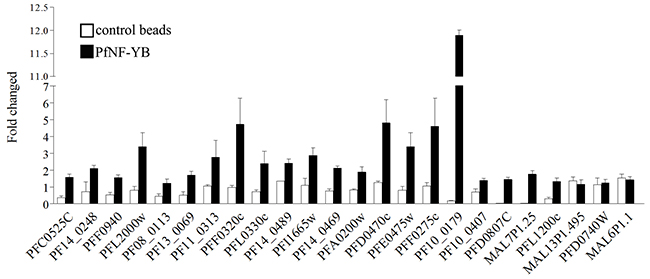
Figure 3: ChIP-on-chip validation. Target genes were selected randomly to perform ChIP/qPCR from schizont stage parasites. Twenty-one different genes showed a PfNF-YB enrichment of at least 2-fold compare to control beads. The last three genes were selected as negative binding targets for PfNF-YB. Input samples were used to normalize the relative values. The control beads were used as a negative control for the ChIP assay. These analyses were derived from three independent assays. The primers used in the analyses can be found in Supplementary Table 3. Gene accession numbers (www.PlasmoDB.org) are indicated at the bottom of the graphic.
PfNF-YB targets are involved in several biological processes in P. falciparum
Analysing the list of identified target genes, we classified and inferred the biological function of the proteins coded by genes bound by PfNF-YB transcription factor using the annotation provided by the PlasmoDB database (version 7.0). From 297 targets identified by the ChIP-on-chip experiment, 42% are of unknown function, 40% are proteins with putative function, 16% are of known function, 1% are pseudogenes and 1% are non-coding RNA (Figure 4A). With respect to the group of 167 proteins of known and putative function associated with a biological process (a gene ontology term), we observed a high diversity of function (Figure 4B). The major groups include proteins involved in RNA translation, protein folding, intracellular transport, cell redox homeostasis and metabolism (for more detail see Supplementary Table 2).
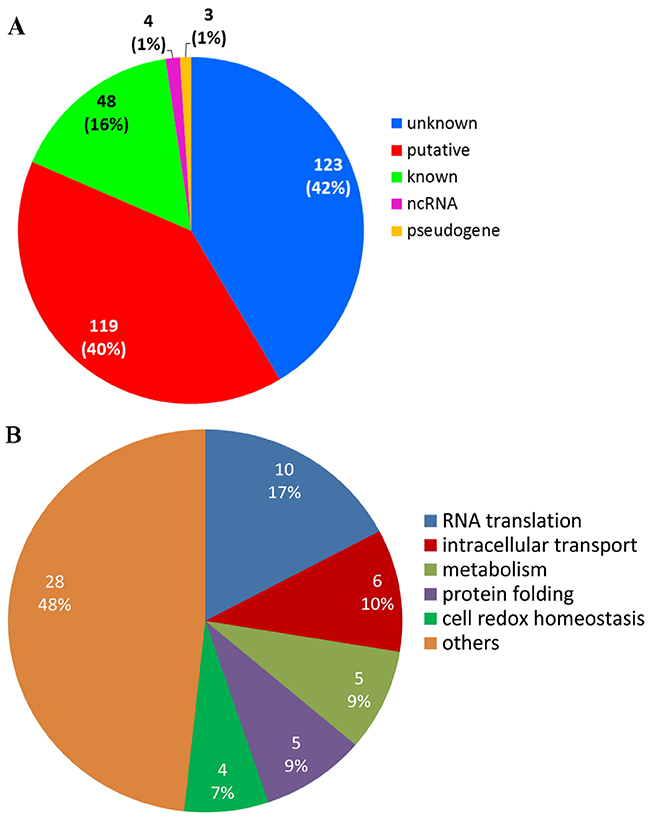
Figure 4: PfNF-YB binds to different gene functions in P. falciparum. (A) PfNF-YB target genes classified by RNA type and protein function. Genes coding for proteins with unknown function (42%) represent the biggest group, followed by the groups represented by the genes with putative (40%) and known (16%) function. (B) Identification of biological processes associated to the target genes bound by PfNF-YB. A group containing 167 coding and putative genes that have a biological process attribute according to the Gene Ontology section from PlasmoDB database were plotted. Fifty-two per cent of these genes are involved in RNA translation, intracellular transport, metabolism, protein folding and cell redox homeostasis.
PfNF-YB recognizes the CCAAT motif in the parasite genome
Transcription factors are crucial in the control of gene expression. Very few transcription factors are known currently in P. falciparum (see review [48]), and this scarcity of information has prevented a better identification of transcription factor binding sites. Using bioinformatics tools, we took advantage of the WebLogo program to elucidate motifs in the promoter regions [4, 18, 49, 50]. In vertebrates, the NF-YB protein family binds to the CCAAT motif, but in P. falciparum the existence and distribution of this motif has not been investigated. Our aims here were to clarify whether the PfNF-YB recognizes the CCAAT motif in P. falciparum and whether or not it binds to others motifs. Probe regions were chosen based on the FDR score and across the range of DNA coverage. Each flanking sequence of the target genes was analysed by ClustalW. First, we searched for the CCAAT motif in both sense and anti-sense strands for the 297 target genes and for the rest of the genome sequence. We found that 157 target genes did not contain the CCAAT motif. Further, we performed Monte Carlo permutation testing [51, 52] in order to determine whether or not the CCAAT motif is over-represented in the putative promoter regions. The simulation took into account the lengths of 297 gene targets identified to obtain a set of new random subsequences from the whole genome, and the number of subsequences that matched at least once with a particular motif was counted. This procedure was repeated 100,000 times to estimate the p-value (i.e., the percentage of repeat executions in which the number of subsequences that contained at least one match equaled or exceeded the actual number of target matches). Indeed, the frequency of the CCAAT motif is significantly higher in the identified PfNF-YB target genes (p-value: 0.00043) than in the rest of the genome. Among these sequences, we found putative promoters of 45 genes containing at least 80% homology in a motif comprising 5 bases upstream and downstream flanking the CCAAT core. We propose 5 consensus sequences for the CCAAT motif in P. falciparum according to the WebLogo analyses (Figure 5A). The P. falciparum genome is the most AT-rich of all the eukaryotes sequenced to date [51, 52] and this high AT content may explain why the nucleotides flanking the CCAAT motif are more AT-rich compared to those in other eukaryotes where NF-Y binds to CCAAT within a more GC-rich sequence environment. In order to better address this situation we analyzed the proportion of A and T nucleotides flanking the CCAAT motif in the PfNF-YB target genes and compared the distribution to the rest of the genome. We examined 5bp both upstream and downstream of the CCAAT motifs in the genome and found that in target genes the frequency of A(T) is on average 5.32(4.88) matches per CCAAT motif, including CCAAT itself which contains two As (one T), whereas in the rest of the genome, the average frequency of A(T) is 5.66 (4.99) per CCAAT motif and flanking sequence. Analyzing each 15 bp-length subsequence throughout the genome, the average A(T) frequency is 6.05 (6.05) per subsequence. Finally, the average number of A's or T's present in the 15 bp subsequences per CCAAT motif in the target genes is 10.2, per CCAAT motif throughout the genome is 10.6, and per 15 bp subsequence throughout the genome is 12.1. These results indicate that the frequencies of A's and T's are not higher in nucleotides flanking CCAAT motifs within PfNF-YB target gene sequences compared to CCAAT motifs generally or compared to 15 bp-subsequences in the rest of the genome. Thus, this analysis indicates that the frequency of A's and T's flanking the CCAAT motif is a general feature of the parasite genome and not a specific sequence preference dictated by the NF-YB motif in the target genes in P. falciparum.
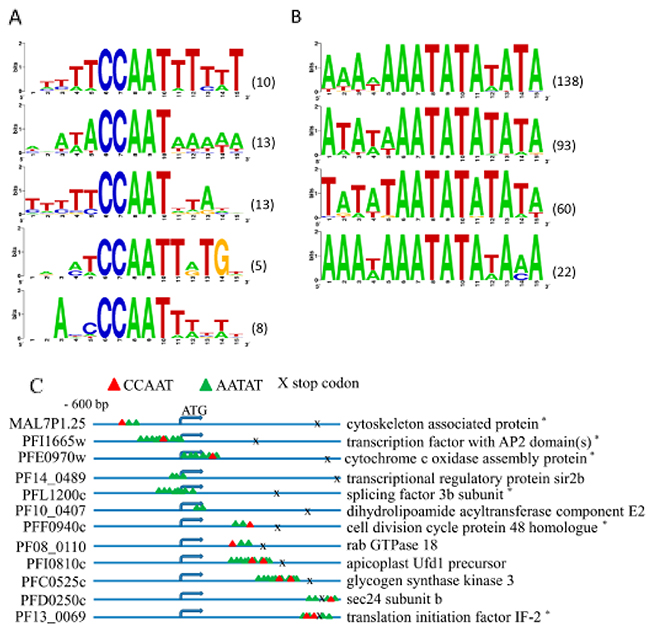
Figure 5: Identification of CCAAT and AATAT motifs bound by PfNF-YB transcription factor. (A) The CCAAT motif sequence recognized by PfNF-YB was found in the promoter regions of 140 genes. Forty-five out of these 140 regions containing 49 CCAAT motifs were investigated for putative consensus sequences. (B) AATAT motif sequence was found in the promoter regions of 297 genes recognized by PfNF-YB. One hundred and twenty-three of these 297 regions containing 313 AATAT motifs were investigated for putative consensus sequences. The probe sequences covering the target genes were selected and assembled, and the putative promoter sequences were analysed by motif finder WebLogo [67]. The graphic shows the relative entropy-based logo of the detected motif. Values in parenthesis represent the number of gene promoters belonging to the consensus motifs. (C) The PfNF-YB target genes present a diversity of binding site regions. The scheme shows the matches to the CCAAT motif and to the putative TATA-box motif. The location of these motifs is both upstream and downstream of the ATG start codon. (*) putative gene.
We also searched for other common motifs using the 297 target genes identified by the ChIP-on-Chip approach. Surprisingly, we discovered several motifs that contained an AATAT core with 5 bases upstream and downstream conserved, which were present in the putative promoters of all 297 genes (Figure 5B). As the AATAT sequence is highly conserved in the binding sites for TATA-box protein, we used the TRANSFAC database to evaluate those sequences that matched TATA box binding sites. Taking into account only the selected sequences, and those clusters with at least four highly similar sequences, we detected 83 putative promoters containing non-TATA box motifs. These sequences were analysed by the WebLogo algorithm and 4 consensus AATAT-based motifs were identified, which may be bound by the PfNF-YB transcription factor, or alternatively the consensus motif for the TATA-box in Plasmodium may exhibit some sequence diversity (Figure 5B). AATAT-based motifs are abundant in plants and are important to control gene expression in Arabidopsis in starvation conditions [53]. This may be a very important finding concerning gene regulation in P. falciparum. The position of the AATAT motif and the CCAAT binding sites in representative genes are illustrated in Figure 5C, displaying all the possible positions of CCAAT and AATAT motifs in the PfNF-YB target genes.
PfNF-YB binds to new motifs
In order to establish whether PfNF-YB binds to a new motif in the putative promoter regions we searched for new sequences and compared these sequences with motifs already described [18]. The analysis showed the presence of 263 matches for AAATG (described in [18]) and 242 matches for ATTTG (a new motif) (Figure 6). The pentameric motif AAATG is a part of the ATG(G)AAATG motif found in maize and wheat histone H3 and H4 mRNAs that contain poly(A) sequences at the 3′-end [54]. ATG(G)AAATG analogous sequences have been identified in the plants arabidopsis, alfalfa, rice and wheat [55]. The ATTTG motif (a CCAAT-like motif) was described as binding to NF-YB in pro-a1(V) and collagen (col5a1) promoter regions that lack the CCAAT canonical sequence (or the reversed complement, ATTGG). The analysis has revealed the presence of this motif in P. falciparum for the first time. Statistical analysis was performed to determine the permutations of motifs located in the same putative promoter regions. Combining the CCAAT, AAATG and ATTTG motifs, we determined the association between them. In Figure 6 only the associations in which the presence of the motif is not due to a random event are displayed. Furthermore, sequences with associations between the three motifs comprise 120 genes (around 40%) out of the 297 identified PFNF-YB target genes. It is important to note that there are twelve target genes lacking any of the three motifs analysed in this study. These results identified AAATG [18] and revealed a new motif ATTTG as a putative PfNF-YB binding sequence, however, whether PFNF-YB binding is direct or indirect was not addressed here.
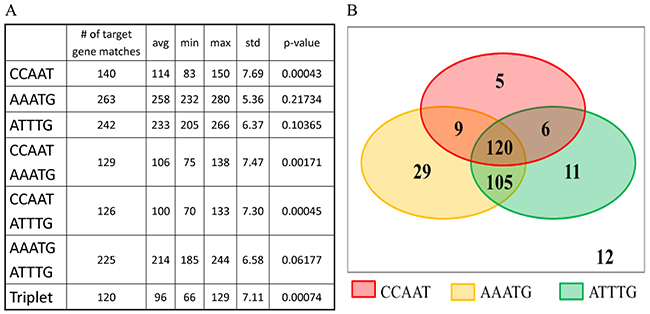
Figure 6: Motif matching in the putative promoter region of PfNF-YB target genes. (A) The Monte Carlo test was performed taking into account the gene target lengths to obtain new random subsequences. This procedure was executed 100,000 times to estimate the p-value (percentage of executions in which the number of subsequences that matched the considered motifs equalled or exceeded the actual number of target matches). The frequency of the CCAAT motif is significantly higher in predicted genes (p-value: 0.00043) than in the rest of the genome. The three motifs CCAAT, AAATG and ATTTG are also significantly more frequent in the PfNF-YB target genes group compared to the rest of the genome (p-value: 0.00074), which suggests that these motifs are strongly associated. (B) A Venn diagram to better illustrate the distribution of the three motifs among the 297 predicted genes. There are 140 matches for CCAAT, 263 matches for AAATG and 242 matches for AATTG. 120 predicted genes (40.4%) match with the three CCAAT, AAATG and ATTTG motifs. Note that only twelve genes (4%) contain none of the three motifs.
DISCUSSION
Our results showed that mRNA PfNF-YB expression is highest in schizont stages in agreement with data from PlasmoDB, and suggesting that it plays an important role at this multiplicative stage of the asexual blood cycle. Therefore, to explore the PfNF-YB role in the asexual cycle of the parasite using chromatin immunoprecipitation (ChiP) was important to elucidate its binding gene target. ChIP is one of the most powerful tools to elucidate protein-DNA interaction and has been applied to study histone- or transcription factor-DNA interaction in Plasmodium [4, 56-59]. In our results we could observe that the noise was a little bit high, but this can be explained by the fact the NF-Y binding to cis-elements containing the highly conserved core sequence CCAAT [60] and rich in the Plasmodium genome. Nevertheless, reports suggest that ~7–8% of human promoters have functional CCAAT motifs [61]. Our results showed that 297 genes were found to bind to PfNF-YB. Considering that the P. falciparum encodes about 5300 genes, we can say that about 5.6% of putative promoters have motif to PfNF-YB binding in the P. falciparum genome. Taking advantage of the P. falciparum genome database, we performed ChIP assays combined with a tiling array and identified, for the first time, several target genes bound by the PfNF-YB transcription factor.
Among genes with putative and known function from ChIP-on-chip results and based on the biological function designation available in PlasmoDB database, we found that PfNF-YB target genes play a role in RNA translation, nucleic acid metabolism, protein translation and transport (Figure 5B and Supplementary Table 2). Genes involved in purine nucleotide metabolism are adenosine monophosphate deaminase (MAL13P1.146) and hypoxanthine phosphoribosyltransferase (Pf10_01221). Moreover, PfNF-YB target gene such as replication protein A large subunit (PFD0470c) was described to participate in DNA replication, repair, recombination and checkpoint processes in other eukaryotic cells [62, 63]. The role of NF-Y in nucleic acid metabolism in mammalian cells is clear and well defined [64]. Considering, a) the increase in PfNF-YB mRNA expression during the differentiation of trophozoites to schizonts (Figure 1) when DNA replication begins in the intraerythrocytic parasite suggests a role for PfNF-YB in DNA duplication. Indeed, DNA synthesis is first observed when the average age of parasites was between 30.5 and 32.5 h and then increase logarithmically until the average age of the parasites are between 44 and 48h [64]; b) the nuclear localization of PfNF-YB in these stages [34] ; c) the in vivo promoter occupancy by PfNF-YB at schizont form (Supplementary Figure 1); and d) its interaction with genes involved in nucleic acid metabolism (Supplementary Table 2), we can hypothesize that PfNF-YB is implicated in DNA replication and multiplication in schizont stage (44 h).
Protein translation and modification processes are among the most promising areas to target for the development of new anti-malarial drugs. PfNF-YB binds to the genes for several translational proteins, for example translation initiation factor IF-2 (PF13_0069) and peptide release factor 1 (PF14_0265). Moreover, we also identified the eukaryotic translation initiation factor 3 subunit 8 (PFL0310c) and several 40S and 60S ribosomal proteins. The third class of gene that PfNF-YB binds is related to transport. Our results demonstrate that PfNF-YB binds to promoter regions of genes for 15 transporter proteins, among them Sec24b (PFD0250c), Rab GTPase 18 (Pf08_0110) and PfMDR2 (Pf14_0455).
Besides the molecular characterization of the transcription factor, it is very important to know the type of motif it binds. In order to infer in silico transcription factor binding sites we applied bioinformatics approaches, such as WebLogo (http://WebLogo.berkeley.edu/). Logos are commonly used for the representation of transcription factor binding site preferences [65]. It also allows us to predict the binding site using the interaction energy with the transcription factor [66, 67]. Therefore, we used WebLogo to determine the motif bound by PfNF-YB in the 297 target genes. The results show that P. falciparum NF-YB binds to a variety of motifs. Although the NF-Y transcription factor binds to the cis-element CCAAT motif in most eukaryotic cells, in P. falciparum we showed by in silico analysis that PfNF-YB does not bind exclusively to this cis-element, since 157 genes do not possess the CCAAT binding site in the region immunoprecipitated with PfNF-YB antibody. ChIP-on-chip analysis shows the interaction between protein and DNA, but does not address whether this interaction is directed or not. Therefore the 157 target genes lacking a CCAAT binding site may be recognized by PfNF-YB due to interactions with other transcription factors.
Our analysis showed the presence of the pentamer motif AAATG (263 matches, Figure 6), corresponding to the well-known Myc transcription factor binding site, which is found in a number of cold-responsive gene promoters in Arabidopsis thaliana [54]. We also detected 242 matches for the ATTTG motif (a CCAAT-like motif), which was described as binding to NF-YB in pro-a1(V) and collagen (col5a1) promoter regions that lack the CCAAT canonical sequence (or the reversed complement, ATTGG), [68]. Based on this information and our results, it appears that the ATTTG motif can replace the CCAAT motif and promote the PfNF-YB binding to DNA in P. falciparum. Furthermore, the WebLogo analysis identified the cis-element AATAT (313 matches) that was identified by in silico TRANSFAC database search as a TATA-box. The presence of a TATA-box motif corroborates the binding of the PfNF-YB transcription factor to putative promoter regions of target genes. The AATAT motif is also associated with gene signaling in inorganic phosphate starvation in Arabidopsis [53]. It is not surprising to find Arabidopsis cis-element sequences in the P. falciparum genome, since the malaria parasite genome is more similar to that of Arabidopsis than other non-Apicomplexan taxa [68].
Recently several works have shown the importance of AP2 genes in controlling blood stage and sexual developmental cycle of Plasmodium. It has been postulated that ap2-g and ap2-g2 proteins play a role as gene repressors during the life cycle progression of P. berghei parasites and they are crucial for gametocytogenesis [69, 70]. By contrast, only non-essential functions were described for ap2-o and ap2-sp proteins in blood stages. Comparing our data with those from Modrzynska and collaborators [69], both for schizonts, we found a total of 50 genes that are potential targets of ap2-g and PfNF-YB proteins (Supplementary Table 4). Out of these 40 (80%) were upregulated after knocking out ap2-g gene in schizont cultures. In addition, the expression of 20 other target genes was also regulated by both ap2-g2 and PfNF-YB proteins. Fifteen (75%) of these genes were upregulated in ap2-g2 knockout parasites. These results suggest that PfNF-YB together with ap2-g or ap2-g2 proteins act as a complex or, at least, separately on different binding sites, to repress the expression of gametocyte genes during the schizont stage. As demonstrated by the authors, gametocytogenesis was lost or significantly reduced in schizonts lacking ap2 or ap2-g2 proteins. Therefore, a hypothetical synergism between ap2-g or ap2-g2 proteins and the PfNF-YB transcription factor, most likely with other regulators, could contribute to these effects on the development of gametocytes in both P. bergei and P. falciparum species. Examining our data and those recently published by Santos and collaborators (2017) we found that PfAp2-l and PfNF-YB proteins regulate 11 common genes in P. falciparum (Supplementary Table 4). PfAp2-l, likewise PfNF-YB, is expressed at the mid to late schizont, and both bind to target genes, such as cAMP-dependent protein kinase catalytic subunit (PKA-c), which was showed to regulate microneme secretion before invasion [71]. ChIP-seq data revealed that PfAp2-l binds the promoter regions of many different histone genes. Indeed, we found that histone H2B is also a common target of PfNF-YB. Moreover, genes encoding proteins involved in RBC remodeling, i. e. early transcribed membrane protein (ETRAMP5), is also a target of PfAp2-l (Santos et al., 2017) and PfNF-YB. This data suggests that P. falciparum transcription factors AP2 and NF-YB regulate DNA/ chromatin genes and host-cell remodeling.
Based on target gene function, PfNF-YB can control nucleic acid metabolism, protein translation, transport and regulation of gene expression. However, it is important to highlight the fact that 42% of all target genes identified have no known function. We demonstrate here that PfNF-YB binds to a variety of motifs showing a divergence compared to the situation in most eukaryotic cells, where the CCAAT motif alone is required for NF-YB interaction with a promoter region. In addition, previously we showed that proteasome inhibitor, bortezomibe (BTZ), besides blockage of ring and trophozoite development also decreases PfNF-YB expression [34], leading us to speculate whether PfNF-YB plays a role in intraerythrocytic developmental cycle of parasite. Indeed, both the transcript and protein levels of PfNF-YB increase following the parasite maturation, resulting in the stimulation of PfNf-YB expression in schizont and a decrease in PfNF-YB expression in ring stage [32, 34, 42]. Here, the results of CHIP-on-Chip assay corroborate the hypothesis that PfNF-YB plays a role in DNA replication and parasite division once genes involved with these biological activities were found enriched in this assay. These data suggest that PfNF-YB could be an interesting anti-malarial candidate, which could acts mainly in schizont stage.
MATERIALS AND METHODS
Culture of plasmodium falciparum
P. falciparum strain 3D7 parasites were cultured in flasks at 37°C and 5% hematocrit in RPMI 1640 medium supplemented with 10% human plasma, and gassed with 90% N2, 5% O2 and 5% CO2 [72]. For synchronization, high parasitemia parasites cultures at the ring stage were centrifuged for 5 min at 800 x g at room temperature, then the supernatant was discarded and the parasitized erythrocytes were incubated for 5 min with 5% sorbitol solution to remove any residual mature parasites. After incubation, the erythrocytes were centrifuged again, washed twice with complete RPMI medium and placed back in culture as previously described [73].
RNA extraction, cDNA synthesis and real-time PCR
Samples from parasites were harvested at 10, 15, 25, 30, 44 and 48 hr after invasion for qPCR assay. Parasitized erythrocytes were lysed using Trizol LS (Invitrogen) according to the manufacturer's instructions. cDNA synthesis was performed with 300 ng of random primers and 500 ng of total RNA previously treated with DNAse, using the Superscript II Kit (Invitrogen) as described in the manufacturer's instructions. Quantitative analysis of the relative levels of specific transcripts in triplicate samples was assessed by SYBER green quantitative real-time PCR (qPCR) in a 7300 Real Time PCR System (ABI) under the following conditions: 50°C for 2 min, 95°C for 10 min, 40 cycles of 95°C for 15 sec; 55°C for 30 sec; 60°C for 30 sec. The efficiency of amplification for each gene was previously determined using dilutions of cDNA or gDNA. Genes outside the 90-110% range were discarded from the analysis. For each reaction 2 to 8 ng of cDNA, oligonucleotides (800 nM final concentration) in Max Mix PCR buffer (ABI) were used. The mRNA was amplified using the following primers. Pf11_0477: 5′-acaagcgaggctagtgacag-3′, 5′-ttcagatattcggttaatggttc-3′; MAL-18S: 5′-aacacaaggaagtttaaggcaacaa-3′, 5′-gcgtgcagcctagttca-3′. The 18S ribosomal RNA gene was amplified as an endogenous control and used to normalize the quantitation results. The reactions were carried out in triplicate and the expression values shown represent the relative amplification from each cDNA sample compared to the control. The significance of the difference between the relative gene expression levels was determined by Student's t-test using three independent assays.
ChIP-on-chip assay
Schizont stage parasites (44 hour post invasion) were isolated from P. falciparum cultures and on three independent occasions were used in chromatin immunoprecipitation assays. Parasites were released from red blood cells by saponin treatment (0.01% saponin, 1x PBS, 1 mM PMSF) as described above, and crosslinking was performed using 2% formaldehyde at 37°C for 10 minutes. Excess formaldehyde was quenched by addition of glycine, with gentle agitation for 5 minutes at room temperature.. The parasites were then lysed in 5 mM HEPES pH 8.0, 85 mM KCl, 0.5% Triton X-100, plus protease inhibitors for 15 minutes. The nuclear fraction was harvested by centrifugation at 3000 rpm and the pellet was incubated with 500 μl of nuclear lysis buffer (50 mM Tris-HCl, 10 mM EDTA pH 8.0, 1% SDS, and protease inhibitors). The DNA was sheared into 200 to 1000 bp fragments by sonication (10 times for 10 seconds using a Sonics Vibra cell Model), and the fractionated chromatin was visualized on an agarose gel stained with ethidium bromide. One milliliter of the supernatant was pre-cleared with 100 μl of protein A-Sepharose beads, by incubation overnight at 4°C with agitation, then the mixture was centrifuged and the pellet discarded. The clarified solution containing fragmented chromatin was incubated overnight at 4°C with 20 μg anti-PfNF-YB or a nonspecific rabbit IgG attached to beads, or with control beads (no antibody attached). The mixture was centrifuged at 300 rpm for 3 minutes and the pellet together with one tenth of the input supernatant were retained. The pellet was washed three times with immunoprecipitation buffer (50 mM HEPES pH 7.5, 500 mM NaCl, 1% Triton X-100, 0.1% sodium deoxycholate, 1 mM EDTA) and then 6 times with wash buffer (10 mM Tris-HCl pH 8.0, 250 mM LiCl, 0.5% NP-40, 0.5% sodium deoxycholate, 1 mM EDTA). Bound material was uncoupled from the Sepharose by reversing the crosslinking at 65°C overnight. The DNA was then purified using the phenol extraction method and dissolved in 30 μl of sterile distilled water. PCR analysis of the DNA immunoprecipitated with PfNF-YB antibody or a control rabbit IgG, and in the beads alone and input fractions was performed as described [74, 75]. The amplicons from PFI1665w (ApiAP2) (5′TCCTGAACGTTCAGGTAAAA3′ and 5′GCAATTCACCTGTTTTCGCT3′), PF14_0489 (Sir2B) (5′TCGTGTCCCGTTAATTTT3′and 5′GTTGATATGCCAGCACCTGA3′) and 18S ribosomal RNA (5′CCACATCTAAGGAAGGCAGC3′ and 5′CACCAGACTTGCCCTCCAA3′) were generated using the following conditions: 95°C for 10 min, 30 cycles of 95°C for 15 sec, 58°C for 30 sec, and 72°C for 45 sec. The 18S ribosomal RNA amplicon served as a loading control [76]. PCR products were resolved by agarose gel electrophoresis and DNA was visualized under UV illumination. The total ChIP DNA was also amplified prior to hybridization with the microarray platform using the Genomeplex WGA kit from Sigma. The quantity and quality of DNA were verified using the Nanodrop and Bioanalyzer apparatus (see below). Amplified dsDNA was labeled with Cy5- (ChIP) or Cy3- (input) and hybridized to a tiling array based on the annotation provided by the PlasmoDB database (version 7:0). The high-density microarray platform of 720,000 features with 3 characters (3 x 720k) covering the whole P. falciparum genome was produced by Roche NimbleGen (USA). The ChIP samples were hybridized with this full P. falciparum genome and the results were analyzed using the Deva program from Roche. The ChIP-on-chip data were normalized using the Tukey bi-weight function to account for differences between the dyes on the array. Roche NimbleGen scaled the ratios in the.gff files by subtracting the bi-weight mean for the probe log2-ratio values for all features on the array from each log2-ratio value.
Accession numbers
The raw ChIP-on-chip data presented and discussed in this publication were deposited in NCBI's Gene Expression Omnibus and are accessible through GEO Series accession number GSE48894 (http://www.ncbi.nlm.nih.gov/geo/query/acc.cgi?acc=GSE48894).
Bioanalyzer
DNA samples from the chromatin immunoprecipitation assay were dissolved in DNase-free water at a final concentration of 5ng/μl. DNA 7500 lab chips (Agilent Technologies) were loaded with samples as recommended by the manufacturer; the micro channels were filled with 9 μl of gel-dye mix in the appropriate wells and the 5μl marker and the samples in 1μl running buffer were loaded. The samples were gently mixed and then the chips were immediately inserted into the Agilent 2100 Bioanalyzer and processed.
ChIP/qPCR validation
The validation of the ChIP-on-chip assay was performed by qPCR using specific primers (Supplementary Table 3). Samples of the input material and material bound to either PfNF-YB-specific or non-specific rabbit IgG coated beads and control beads were used at the same molar concentration to verify the IP enrichment compared with the bead-alone control. The qPCR was performed using Power SYBR green mix (Applied Biosystems) in triplicate and from three independent experiments. The relative amount of DNA of target genes was normalized against the input samples and the values of the cycle threshold were calculated for the absolute amount of DNA.
Motif analysis
To analyze the promoter regions of putative candidate genes, we used P. falciparum genome sequences available in the databases: http://plasmodb.org/plasmo/ and http://genome.ucsc.edu/. Sequences upstream of the ATG translation initiation codon, were evaluated as potential promoter regions by analyzing 500 bp upstream and downstream of the PFNF-YB binding site. The WebLogo program was used to represent the DNA sequence of all target genes from the ChIP-on-chip assay.
Statistical analyses
The statistical significance of differences between means was analysed by Student's t test or Monte Carlo methods, as indicated. The Monte Carlo permutation test [77, 78] was used to determine whether or not the CCAAT, AAATG, ATTTG and AATAT motifs are overrepresented in the putative promoter regions. The simulation took into account the lengths of the 297 gene targets identified. The number of subsequences that matched at least once with a particular motif was counted and the procedure was repeated 100,000 times to derive a p-value.
Author contributions
Conceived and designed the experiments: WRL, KSP, CRSG. Performed the experiments: WRL, Nimblegene (Roche). Analyzed the data: WRL, DCM-Jr, KSP, PT, RFH, CRSG. Designed the script for ChIP-on-chip data: DCM-Jr. Wrote the paper: All authors read and approved the final manuscript.
ACKNOWLEDGMENTS AND FUNDING
We thank São Paulo Research Foundation (FAPESP) Process 2011/51295-5 for funding C.R.S.G. W.R.L is a FAPESP fellow. FAPESP 2011/50761-2 and 2015/01587-0 for funding DCM-Jr and RFH. Our thanks to Hemocentro Hospital do Servidor Público for providing us blood and plasma. We also thanks NUSCEP (Nucleus for research on cellular signaling on host-pathogen interactions).
COMPETING INTEREST
The authors declare that they have no conflicts of interest.
REFERENCES
1. Garcia CR, de Azevedo MF, Wunderlich G, Budu A, Young JA, Bannister L. Plasmodium in the postgenomic era: new insights into the molecular cell biology of malaria parasites. Int Rev Cell Mol Biol. 2008; 266:85–156.
2. Maier AG, Cooke BM, Cowman AF, Tilley L. Malaria parasite proteins that remodel the host erythrocyte. Nat Rev Microbiol. 2009; 7:341–54.
3. Gardner MJ, Hall N, Fung E, White O, Berriman M, Hyman RW, Carlton JM, Pain A, Nelson KE, Bowman S, Paulsen IT, James K, Eisen JA, et al. Genome sequence of the human malaria parasite Plasmodium falciparum. Nature. 2002; 419:498–511.
4. Campbell TL, De Silva EK, Olszewski KL, Elemento O, Llinás M. Identification and genome-wide prediction of DNA binding specificities for the ApiAP2 family of regulators from the malaria parasite. PLoS Pathog. 2010; 6:e1001165.
5. Painter HJ, Campbell TL, Llinás M. The Apicomplexan AP2 family: integral factors regulating Plasmodium development. Mol Biochem Parasitol. 2011; 176:1–7.
6. Sinha A, Hughes KR, Modrzynska KK, Otto TD, Pfander C, Dickens NJ, Religa AA, Bushell E, Graham AL, Cameron R, Kafsack BF, Williams AE, Llinas M, et al. A cascade of DNA-binding proteins for sexual commitment and development in Plasmodium. Nature. 2014; 507:253–57.
7. Lanzer M, de Bruin D, Wertheimer SP, Ravetch JV. Organization of chromosomes in Plasmodium falciparum: a model for generating karyotypic diversity. Parasitol Today. 1994; 10:114–17.
8. de Bruin D, Lanzer M, Ravetch JV. The polymorphic subtelomeric regions of Plasmodium falciparum chromosomes contain arrays of repetitive sequence elements. Proc Natl Acad Sci USA. 1994; 91:619–23.
9. Alano P, Silvestrini F, Roca L. Structure and polymorphism of the upstream region of the pfg27/25 gene, transcriptionally regulated in gametocytogenesis of Plasmodium falciparum. Mol Biochem Parasitol. 1996; 79:207–17.
10. Cheesman S, Horrocks P, Tosh K, Kilbey B. Intraerythrocytic expression of topoisomerase II from Plasmodium falciparum is developmentally regulated. Mol Biochem Parasitol. 1998; 92:39–46.
11. Scherf A, Hernandez-Rivas R, Buffet P, Bottius E, Benatar C, Pouvelle B, Gysin J, Lanzer M. Antigenic variation in malaria: in situ switching, relaxed and mutually exclusive transcription of var genes during intra-erythrocytic development in Plasmodium falciparum. EMBO J. 1998; 17:5418–26.
12. Horrocks P, Kilbey BJ. Physical and functional mapping of the transcriptional start sites of Plasmodium falciparum proliferating cell nuclear antigen. Mol Biochem Parasitol. 1996; 82:207–15.
13. Coleman BI, Duraisingh MT. Transcriptional control and gene silencing in Plasmodium falciparum. Cell Microbiol. 2008; 10:1935–46.
14. Coulson RM, Hall N, Ouzounis CA. Comparative genomics of transcriptional control in the human malaria parasite Plasmodium falciparum. Genome Res. 2004; 14:1548–54.
15. Callebaut I, Prat K, Meurice E, Mornon JP, Tomavo S. Prediction of the general transcription factors associated with RNA polymerase II in Plasmodium falciparum: conserved features and differences relative to other eukaryotes. BMC Genomics. 2005; 6:100–20.
16. Gissot M, Briquet S, Refour P, Boschet C, Vaquero C. PfMyb1, a Plasmodium falciparum transcription factor, is required for intra-erythrocytic growth and controls key genes for cell cycle regulation. J Mol Biol. 2005; 346:29–42.
17. Yuda M, Iwanaga S, Shigenobu S, Mair GR, Janse CJ, Waters AP, Kato T, Kaneko I. Identification of a transcription factor in the mosquito-invasive stage of malaria parasites. Mol Microbiol. 2009; 71:1402–14.
18. Harris EY, Ponts N, Le Roch KG, Lonardi S. Chromatin-driven de novo discovery of DNA binding motifs in the human malaria parasite. BMC Genomics. 2011; 12:601.
19. Beraldo FH, Mikoshiba K, Garcia CR. Human malarial parasite, Plasmodium falciparum, displays capacitative calcium entry: 2-aminoethyl diphenylborinate blocks the signal transduction pathway of melatonin action on the P. falciparum cell cycle. J Pineal Res. 2007; 43:360–64.
20. Hotta CT, Gazarini ML, Beraldo FH, Varotti FP, Lopes C, Markus RP, Pozzan T, Garcia CR. Calcium-dependent modulation by melatonin of the circadian rhythm in malarial parasites. Nat Cell Biol. 2000; 2:466–68.
21. Alves E, Bartlett PJ, Garcia CR, Thomas AP. Melatonin and IP3-induced Ca2+ release from intracellular stores in the malaria parasite Plasmodium falciparum within infected red blood cells. J Biol Chem. 2011; 286:5905–12.
22. Beraldo FH, Almeida FM, da Silva AM, Garcia CR. Cyclic AMP and calcium interplay as second messengers in melatonin-dependent regulation of Plasmodium falciparum cell cycle. J Cell Biol. 2005; 170:551–57.
23. Berridge MJ. Calcium signalling remodelling and disease. Biochem Soc Trans. 2012; 40:297–309.
24. Merckx A, Bouyer G, Thomas SL, Langsley G, Egée S. Anion channels in Plasmodium-falciparum-infected erythrocytes and protein kinase A. Trends Parasitol. 2009; 25:139–44.
25. Rached FB, Ndjembo-Ezougou C, Chandran S, Talabani H, Yera H, Dandavate V, Bourdoncle P, Meissner M, Tatu U, Langsley G. Construction of a Plasmodium falciparum Rab-interactome identifies CK1 and PKA as Rab-effector kinases in malaria parasites. Biol Cell. 2012; 104:34–47.
26. Koyama FC, Chakrabarti D, Garcia CR. Molecular machinery of signal transduction and cell cycle regulation in Plasmodium. Mol Biochem Parasitol. 2009; 165:1–7.
27. Borrelli E, Montmayeur JP, Foulkes NS, Sassone-Corsi P. Signal transduction and gene control: the cAMP pathway. Crit Rev Oncog. 1992; 3:321–38.
28. Ha E, Han E, Park HJ, Kim HJ, Hong MS, Hong SJ, Yoon KS, Kang I, Cho YH, Chung JH, Yim SV, Baik HH. Microarray analysis of transcription factor gene expression in melatonin-treated human peripheral blood mononuclear cells. J Pineal Res. 2006; 40:305–11.
29. Lalli E, Sassone-Corsi P. Signal transduction and gene regulation: the nuclear response to cAMP. J Biol Chem. 1994; 269:17359–62.
30. Ziff EB. Transcription factors: a new family gathers at the cAMP response site. Trends Genet. 1990; 6:69–72.
31. Barbado M, Fablet K, Ronjat M, De Waard M. Gene regulation by voltage-dependent calcium channels. Biochim Biophys Acta. 2009; 1793:1096–104.
32. Silvestrini F, Bozdech Z, Lanfrancotti A, Di Giulio E, Bultrini E, Picci L, Derisi JL, Pizzi E, Alano P. Genome-wide identification of genes upregulated at the onset of gametocytogenesis in Plasmodium falciparum. Mol Biochem Parasitol. 2005; 143:100–10.
33. Currà C, Gessmann R, Pace T, Picci L, Peruzzi G, Varamogianni-Mamatsi V, Spanos L, Garcia CR, Spaccapelo R, Ponzi M, Siden-Kiamos I. Release of Plasmodium sporozoites requires proteins with histone-fold dimerization domains. Nat Commun. 2016; 7:13846.
34. Lima WR, Moraes M, Alves E, Azevedo MF, Passos DO, Garcia CR. The PfNF-YB transcription factor is a downstream target of melatonin and cAMP signalling in the human malaria parasite Plasmodium falciparum. J Pineal Res. 2013; 54:145–53.
35. Dorn A, Bollekens J, Staub A, Benoist C, Mathis D. A multiplicity of CCAAT box-binding proteins. Cell. 1987; 50:863–72.
36. Hatamochi A, Golumbek PT, Van Schaftingen E, de Crombrugghe B. A CCAAT DNA binding factor consisting of two different components that are both required for DNA binding. J Biol Chem. 1988; 263:5940–47.
37. Hooft van Huijsduijnen RA, Bollekens J, Dorn A, Benoist C, Mathis D. Properties of a CCAAT box-binding protein. Nucleic Acids Res. 1987; 15:7265–82.
38. Hu Q, Bhattacharya C, Maity SN. CCAAT binding factor (CBF) binding mediates cell cycle activation of topoisomerase iialpha. conventional CBF activation domains are not required. J Biol Chem. United States; 2002; 277:37191–200.
39. Tanimoto A, Chen H, Kao CY, Moran E, Sasaguri Y, Padmanabhan R. Transactivation of the human cdc2 promoter by adenovirus E1A in cycling cells is mediated by induction of a 110-kDa CCAAT-box-binding factor. Oncogene. 1998; 17:3103–14.
40. Maity SN, de Crombrugghe B. Role of the CCAAT-binding protein CBF/NF-Y in transcription. Trends Biochem Sci. 1998; 23:174–78.
41. Calderwood MS, Gannoun-Zaki L, Wellems TE, Deitsch KW. Plasmodium falciparum var genes are regulated by two regions with separate promoters, one upstream of the coding region and a second within the intron. J Biol Chem. 2003; 278:34125–32.
42. Bozdech Z, Llinás M, Pulliam BL, Wong ED, Zhu J, DeRisi JL. The transcriptome of the intraerythrocytic developmental cycle of Plasmodium falciparum. PLoS Biol. 2003; 1:E5.
43. Gannoun-Zaki L, Jost A, Mu J, Deitsch KW, Wellems TE. A silenced Plasmodium falciparum var promoter can be activated in vivo through spontaneous deletion of a silencing element in the intron. Eukaryot Cell. 2005; 4:490–92.
44. Brick K, Watanabe J, Pizzi E. Core promoters are predicted by their distinct physicochemical properties in the genome of Plasmodium falciparum. Genome Biol. 2008; 9:R178.
45. Scholtmeijer K, Wösten HA, Springer J, Wessels JG. Effect of introns and AT-rich sequences on expression of the bacterial hygromycin B resistance gene in the basidiomycete Schizophyllum commune. Appl Environ Microbiol. 2001; 67:481–83.
46. Schuren FH, Wessels JG. Expression of heterologous genes in Schizophyllum commune is often hampered by the formation of truncated transcripts. Curr Genet. 1998; 33:151–56.
47. Tuteja R. Helicases involved in splicing from malaria parasite Plasmodium falciparum. Parasitol Int. 2011; 60:335–40.
48. De Silva EK, Gehrke AR, Olszewski K, León I, Chahal JS, Bulyk ML, Llinás M. Specific DNA-binding by apicomplexan AP2 transcription factors. Proc Natl Acad Sci USA. 2008; 105:8393–98.
49. Llinás M, Deitsch KW, Voss TS. Plasmodium gene regulation: far more to factor in. Trends Parasitol. 2008; 24:551–56.
50. Dwass M. Modified randomization tests for nonparametric hypotheses. Ann Math Stat. 1957; 28:181–87.
51. Nichols TE, Holmes AP. Nonparametric permutation tests for functional neuroimaging: a primer with examples. Hum Brain Mapp. 2002; 15:1–25.
52. Chen A, Gu M, Sun S, Zhu L, Hong S, Xu G. Identification of two conserved cis-acting elements, MYCS and P1BS, involved in the regulation of mycorrhiza-activated phosphate transporters in eudicot species. New Phytol. 2011; 189:1157–69.
53. Abe H, Yamaguchi-Shinozaki K, Urao T, Iwasaki T, Hosokawa D, Shinozaki K. Role of arabidopsis MYC and MYB homologs in drought- and abscisic acid-regulated gene expression. Plant Cell. 1997; 9:1859–68.
54. Ohtsubo N, Iwabuchi M. The conserved 3’-flanking sequence, AATGGAAATG, of the wheat histone H3 gene is necessary for the accurate 3’-end formation of mRNA. Nucleic Acids Res. 1994; 22:1052–58.
55. Salcedo-Amaya AM, van Driel MA, Alako BT, Trelle MB, van den Elzen AM, Cohen AM, Janssen-Megens EM, van de Vegte-Bolmer M, Selzer RR, Iniguez AL, Green RD, Sauerwein RW, Jensen ON, Stunnenberg HG. Dynamic histone H3 epigenome marking during the intraerythrocytic cycle of Plasmodium falciparum. Proc Natl Acad Sci USA. 2009; 106:9655–60.
56. Flueck C, Bartfai R, Volz J, Niederwieser I, Salcedo-Amaya AM, Alako BT, Ehlgen F, Ralph SA, Cowman AF, Bozdech Z, Stunnenberg HG, Voss TS. Plasmodium falciparum heterochromatin protein 1 marks genomic loci linked to phenotypic variation of exported virulence factors. PLoS Pathog. 2009; 5:e1000569.
57. Lopez-Rubio JJ, Mancio-Silva L, Scherf A. Genome-wide analysis of heterochromatin associates clonally variant gene regulation with perinuclear repressive centers in malaria parasites. Cell Host Microbe. 2009; 5:179–90.
58. Mair GR, Braks JA, Garver LS, Wiegant JC, Hall N, Dirks RW, Khan SM, Dimopoulos G, Janse CJ, Waters AP. Regulation of sexual development of Plasmodium by translational repression. Science. 2006; 313:667–69.
59. Cao S, Kumimoto RW, Siriwardana CL, Risinger JR, Holt BF 3rd. Identification and characterization of NF-Y transcription factor families in the monocot model plant Brachypodium distachyon. PLoS One. 2011; 6:e21805.
60. FitzGerald PC, Shlyakhtenko A, Mir AA, Vinson C. Clustering of DNA sequences in human promoters. Genome Res. 2004; 14:1562–74.
61. Brush GS, Kelly TJ. Phosphorylation of the replication protein A large subunit in the Saccharomyces cerevisiae checkpoint response. Nucleic Acids Res. 2000; 28:3725–32.
62. Wold MS. Replication protein A: a heterotrimeric, single-stranded DNA-binding protein required for eukaryotic DNA metabolism. Annu Rev Biochem. 1997; 66:61–92.
63. Dolfini D, Gatta R, Mantovani R. NF-Y and the transcriptional activation of CCAAT promoters. Crit Rev Biochem Mol Biol. 2012; 47:29–49.
64. Inselburg J, Banyal HS. Synthesis of DNA during the asexual cycle of Plasmodium falciparum in culture. Mol Biochem Parasitol. 1984; 10:79–87.
65. Takeda Y, Sarait A, Rivera VM. Analysis of the sequence-specific interactions between cro repressor and operator DNA by systematic base substitution experiments (DNA-protein interaction/DNA sequence recognition/additivity of binding free energy changes). Biochemistry. 1989; 86:439–43.
66. Man TK, Stormo GD. Non-independence of Mnt repressor-operator interaction determined by a new quantitative multiple fluorescence relative affinity (QuMFRA) assay. Nucleic Acids Res. 2001; 29:2471–78.
67. Sakata-Takatani K, Matsuo N, Sumiyoshi H, Tsuda T, Yoshioka H. Identification of a functional CBF-binding CCAAT-like motif in the core promoter of the mouse pro-alpha 1(V) collagen gene (Col5a1). Matrix Biol. 2004; 23:87–99.
68. Bastien O, Lespinats S, Roy S, Métayer K, Fertil B, Codani JJ, Maréchal E. Analysis of the compositional biases in Plasmodium falciparum genome and proteome using Arabidopsis thaliana as a reference. Gene. 2004; 336:163–73.
69. Modrzynska K, Pfander C, Chappell L, Yu L, Suarez C, Dundas K, Gomes AR, Goulding D, Rayner JC, Choudhary J, Billker O. A knockout screen of ApiAP2 genes reveals networks of interacting transcriptional regulators controlling the Plasmodium life cycle. Cell Host Microbe. 2017; 21:11–22.
70. Yuda M, Iwanaga S, Kaneko I, Kato T. Global transcriptional repression: an initial and essential step for Plasmodium sexual development. Proc Natl Acad Sci USA. 2015; 112:12824–29.
71. Dawn A, Singh S, More KR, Siddiqui FA, Pachikara N, Ramdani G, Langsley G, Chitnis CE. The central role of cAMP in regulating Plasmodium falciparum merozoite invasion of human erythrocytes. PLoS Pathog. 2014; 10:e1004520.
72. Trager W, Jensen JB. Human malaria parasites in continuous culture. Science. 1976; 193:673–75.
73. Lambros C, Vanderberg JP. Synchronization of Plasmodium falciparum erythrocytic stages in culture. J Parasitol. 1979; 65:418–20.
74. Papadopoulou K, Chen JS, Mead E, Feoktistova A, Petit C, Agarwal M, Jamal M, Malik A, Spanos A, Sedgwick SG, Karagiannis J, Balasubramanian MK, Gould KL, McInerny CJ. Regulation of cell cycle-specific gene expression in fission yeast by the Cdc14p-like phosphatase Clp1p. J Cell Sci. 2010; 123:4374–81.
75. Peterson LF, Boyapati A, Ranganathan V, Iwama A, Tenen DG, Tsai S, Zhang DE. The hematopoietic transcription factor AML1 (RUNX1) is negatively regulated by the cell cycle protein cyclin D3. Mol Cell Biol. 2005; 25:10205–19.
76. Lima WR, Parreira KS, Devuyst O, Caplanusi A, N’kuli F, Marien B, Van Der Smissen P, Alves PM, Verroust P, Christensen EI, Terzi F, Matter K, Balda MS, et al. ZONAB promotes proliferation and represses differentiation of proximal tubule epithelial cells. J Am Soc Nephrol. 2010; 21:478–88.
77. Kalos MH, Whitlock PA. Basics. Monte carlo methods. New York, NY, USA: Wiley-Interscience; 1986: 1.
78. Metropolis N, Ulam S. The Monte Carlo method. J Am Stat Assoc. 1949; 44:335–41.