INTRODUCTION
Liver receptor homologue 1 (LRH-1, NR5A2) is a nuclear receptor and acts as a transcription factor with roles in embryogenesis, steroid and cholesterol metabolism, inflammation, and in several cancers, including gastrointestinal malignancies and breast cancer [1–9]. In breast cancer cell lines, multiple studies have shown that LRH-1 has tumour-promoting roles in estrogen production, estrogen receptor (ER) signalling, cell cycle control, and cellular migration and invasion [1, 7, 8, 10–15]. LRH-1 also stimulates local oestrogen production by upregulating aromatase activity in breast adipose tissue [10, 12, 13] and by enhancing the effect of prostaglandin E2 on aromatase expression [12]. In addition, LRH-1 and ER each promote the expression of the other [11, 15], share many binding sites [14], and co-operatively regulate the expression of ER target genes [11, 14].
LRH-1 shows ER- independent actions too, such as modulating expression of genes involved in cell cycle control including upregulating the expression of CCND1 [7], MYC [7] and BCL2 [7], and suppressing CDKN1A expression [1]. Reduction of cell proliferation upon LRH-1 knockdown occurs in a p53-independent manner [1] and results in an increased proportion of cells in the G0/G1 phase of the cell cycle and reduction of cells in the S and G2/M phases [15]. Compared with ER-positive breast cancer cells, the anti-proliferative effect of LRH-1 knockdown on the cell cycle is more pronounced in MCF-7 in the absence of E2 [15], in MCF7- derived anti-estrogen-resistant cell lines (MCF7/LCC2 and MCF7/LCC9) and in the ER-negative cell line BT-549 [1, 7]. This suggests that LRH-1 may have a greater role in driving cell proliferation in breast cancer cells in the absence of functional ER, perhaps by providing an alternative mechanism for regulation of ER target genes. Indeed, higher LRH-1 expression is present in MCF7/LCC2 and MCF7/LCC9 cell lines compared with parental MCF7 cells [7], and overexpression of LRH-1 in the ER-negative cell line MDA-MB-231 results in significant up-regulation of the ER target gene GREB1 [11].
Although LRH-1 mediates processes that promote tumorigenesis in both estrogen-driven and estrogen-independent breast cancer cells, the direct role of LRH-1 in human breast cancer remains unexplored. Most LRH-1 studies have been performed on breast cancer cell lines and data from breast cancer tissues is limited, both regarding LRH-1 expression and the relationship of LRH-1 with tumour biology. Moreover, although LRH-1 expression is influenced by ER in ER-positive breast cancer, very little is known about alternative mechanisms controlling LRH-1 expression, in particular how it is regulated in ER-negative breast tumours, and in ER-positive breast cancers resistant to anti-estrogenic therapy.
LRH-1 is encoded by the NR5A2 gene which is located on chromosome 1 at band q32.1. There are at least five described LRH-1 mRNA transcripts [2, 16–20], generated by different transcription initiation sites as well as alternative splicing, four of which are associated with protein products [21] (Table 1, Figure 1). Regulation of these transcripts may be controlled by methylation, as six CpG islands are present in the gene region. A 501 amino acid protein, first described and named variant 4 by Thiruchelvam et al. in 2011 [20], is reported to be the predominant mRNA variant in breast cancer cell lines and to be highly estrogen regulated compared with other variants [20]. Therefore the aims of this study are to assess the importance of LRH-1 in situ and invasive breast cancer, in particular to investigate (1) NR5A2 transcript expression in invasive breast cancers (2) the role of DNA methylation in regulating the expression of NR5A2, (3) the level and pattern of expression of LRH-1 protein in a cohort of ductal carcinoma in situ (DCIS) and invasive breast carcinomas to assess its potential role in tumour progression and (4) the relationship between LRH-1 expression and clinicopathological features.
Table 1: (A) NR5A2 transcripts and (B) CpG islands
A. NR5A2 transcripts
Genomic sequence (GRCh37/hg19) |
Number of exons |
RefSeq accession |
RNA accession |
Protein length |
|---|---|---|---|---|
chr1:199,996,730-200,008,923 |
3 |
NM_205860 |
AF124248.1 |
No protein |
chr1:199,996,730-200,146,550 |
8 |
NM_205860 |
NM_205860.2 |
541aa |
chr1:199,996,730-200,146,550 |
7 |
NM_003822 |
NM_003822.4 |
495aa |
chr1:200,008,658-200,146,550 |
7 |
NM_001276464 (“variant 4”) |
AK304344.1 (“variant 4”) |
501aa |
chr1:200,011,953-200,146,550 |
6 |
NM_001276464 |
NM_001276464.1 |
469aa |
B. CpG islands within NR5A2
CpG island |
Number of CpG dinucleotides |
|
|---|---|---|
1 |
chr1:200,004,475-200,004,933 |
36 |
2 |
chr1:200,008,393-200,009,047 |
72 |
3 |
chr1:200,009,808-200,010,036 |
24 |
4 |
chr1:200,010,626-200,010,832 |
16 |
5 |
chr1:200,011,401-200,012,055 |
55 |
6 |
chr1:200,116,697-200,117,204 |
45 |
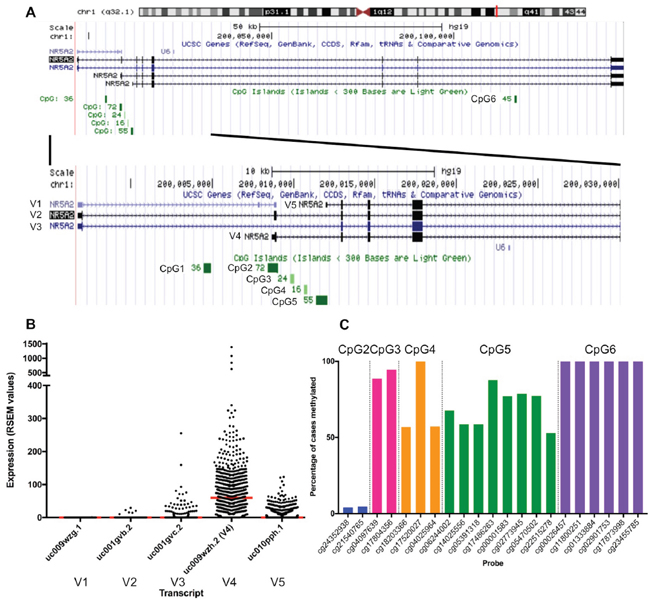
Figure 1: (A) Genomic region of NR5A2 showing the five transcripts (V1-V5) and CpG islands (CpG1 – CpG6). (B) mRNA expression of NR5A2 transcripts in TCGA invasive breast cancer cohort. RSEM, RNA-Seq by Expectation Maximization. Red line indicates median. (C) Methylation frequencies of HM450 probes within NR5A2 CpG islands CpG2-CpG6 in TCGA invasive breast cancer cohort.
RESULTS
NR5A2 mRNA expression and its relationship to DNA methylation using The Cancer Genome Atlas (TCGA) data
Six CpG islands are located within the NR5A2 gene [16, 22] (Table 1, Figure 1A) with several CpG islands associated with variant 4. The second CpG island (CpG2) of these six is situated immediately upstream of variant 4 and four intragenic CpG islands (third, fourth, fifth and sixth CpG islands - CpG3, CpG4, CpG5 and CpG6, respectively) are located nearby, suggesting that methylation of these islands may have a role in regulating expression of variant 4. The first CpG island (CpG1) is located approximately 4 kb upstream of variant 4.
Isoform-specific expression data and methylation data was available for 756 samples. ER, PR, and HER2 status were available for 371, 333, and 468 cases, respectively. The majority of cases were ER positive (86.8%, 322/371), PR positive (70.6%, 235/333), and HER2 negative (80.8%, 378/468). No information regarding tumour grade was available.
Similar to published breast cancer cell line data [15, 20, 23], variant 4 was the predominantly expressed transcript in invasive breast cancers in the TCGA cohort (Figure 1). Variant 4 expression levels were significantly higher in ER-positive tumours compared with ER-negative tumours (ER positive mean 89.6 ± 116.6 RNA-Seq by Expectation Maximization (RSEM) values; ER negative mean 65.7 ± 64.9 RSEM values, p=0.011). This is not a consequence of the frequent copy number gain of NR5A2 in ER-positive tumours, as copy number and mRNA expression are not positively correlated in TCGA data (Pearson r=-0.093). Variant 4 expression did not differ significantly based on PR or HER2 status (p=0.101 and p=0.079 respectively).
CpG6 was universally methylated at all six probe sites. CpG2 was the least frequently methylated, with just 4.9% (40/756) of cases methylated for both CpG2 probes. CpG3, CpG4, and CpG5 showed some degree of methylation in the vast majority of cases (methylation present in 95.2% (720/756), 100%, and 95.8% (724/756) of cases, respectively), however methylation was somewhat heterogeneous (all probes within CpG island methylated in 88.0% (665/756), 48.1% (364/756), 31.6% (239/756) of cases, respectively) (Figure 1).
The expression of variant 4 was significantly lower in the 28 cases methylated at both CpG2 probes compared with cases not showing this methylation profile (p=0.036, Figure 2). In contrast, cases methylated at both CpG3 probes (n= 665) showed higher variant 4 expression compared with cases unmethylated at one or both probes (p=0.01, Figure 2). All cases methylated at both CpG2 probes were also methylated at both CpG3 probes, while 95.8% (637/665) of cases methylated at both CpG3 sites were not fully methylated at CpG2. The methylation status of CpG4 and CpG5 were not significantly associated with variant 4 expression.
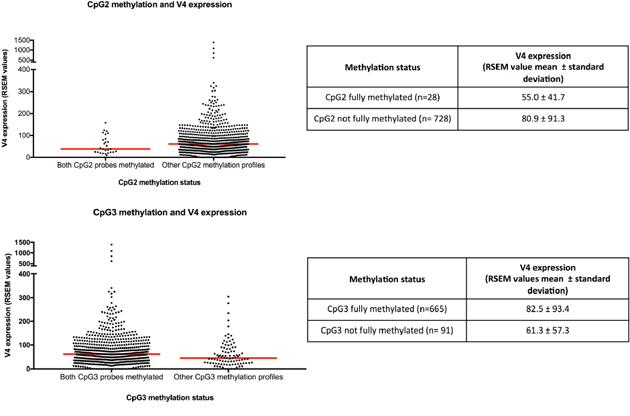
Figure 2: CpG2 and CpG3 methylation and NR5A2 variant 4 expression. RSEM, RNA-Seq by Expectation Maximization. Red line indicates median.
LRH-1 protein expression
While NR5A2 mRNA was associated with ER status in invasive breast cancer, post-translational regulation of LRH-1 protein may affect such associations with breast cancer characteristics. Thus we evaluated LRH-1 protein using immunohistochemistry (IHC), successfully obtaining results from 329 cases of breast cancer, comprising 175 DCIS (146 pure DCIS and 29 DCIS occurring synchronously with invasive carcinoma) and 154 invasive carcinomas. When present, IHC nuclear reactivity was observed in all epithelial cells throughout the tumours, with four patterns of nuclear positivity observed (Figure 3). Nuclear positivity showed a finely dispersed pattern (pattern 1), a sparse punctate pattern (pattern 2), a dense punctate pattern (pattern 2+), and a coarse granular pattern (pattern 3). Patterns 2, 2+, and 3 also showed finely dispersed nuclear positivity in the background. Most cases showed uniformity of nuclear pattern; where this was not the case the dominant pattern was recorded. 1.2% of cases (4/329) showed no nuclear positivity. The frequencies of patterns 1, 2, 2+, and 3 staining were 28.3% (93/329), 50.5% (166/329), 14.0% (46/329), and 6.1% (20/329), respectively. Cases with no IHC staining or pattern 1 or 2 staining were considered to be ‘IHC fine’. Patterns 2+ and 3 were considered to be ‘IHC granular’.
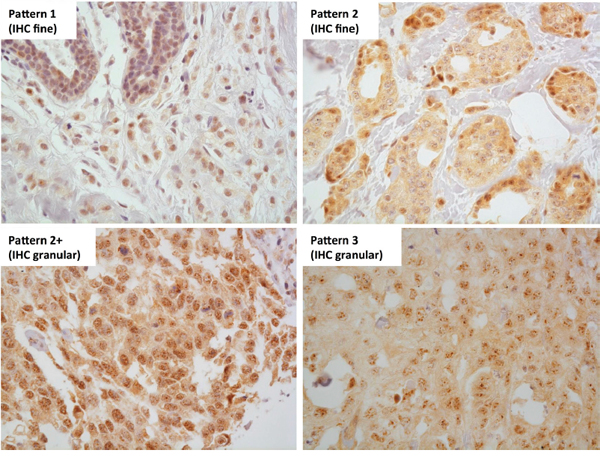
Figure 3: LRH-1 IHC nuclear reactivity patterns. Four patterns of LRH-1 IHC nuclear reactivity were observed - a finely granular pattern, similar to that seen in normal breast epithelium (pattern 1), a sparse punctate pattern (pattern 2), a dense punctate pattern (pattern 2+), and a coarse granular pattern (pattern 3). Patterns 1 and 2 were classified as LRH-1 IHC fine and patterns 2+ and 3 were classified as LRH-1 IHC granular. 400x magnification.
There was no difference in LRH-1 staining between DCIS and invasive carcinoma (p=0.411, Table 2). In the overall cohort (Table 2), the IHC-granular group was significantly associated with high grade (p<0.0005), ER negativity (p=0.008), PR negativity (p=0.003), HER2 amplification (p=0.034) and non-luminal intrinsic subtypes (p=0.018) compared with cases not showing this pattern. There was a trend for LRH-1 granular tumours to be larger but this was not statistically significant (p=0.054). There was no association with age or lymph node status. When separating out by tumour type, invasive tumours showing IHC granular staining were significantly correlated with high grade (p<0.0005), ER negativity (p=0.038), PR negativity (p=0.013), and non-luminal intrinsic subtypes (p=0.038) (Table 2). In DCIS, IHC-granular staining patterns were associated with high nuclear grade (p=0.033) (Table 2).
Table 2: LRH-1 IHC and tumour phenotype
Overall cohort (n=329) |
Invasive carcinoma (n=154) |
DCIS (n=175) |
|||||||
|---|---|---|---|---|---|---|---|---|---|
IHC granular |
IHC fine |
p-value |
IHC granular |
IHC fine |
p-value |
IHC granular |
IHC fine |
p-value |
|
Number |
66 |
263 |
34 |
120 |
32 |
143 |
|||
Tumour type |
|||||||||
Invasive |
34 |
120 |
0.411 |
N/A |
N/A |
N/A |
N/A |
N/A |
N/A |
DCIS |
32 |
143 |
|||||||
DCIS type |
|||||||||
Pure |
N/A |
N/A |
N/A |
N/A |
N/A |
N/A |
29 |
117 |
0.298 |
Mixed |
3 |
26 |
|||||||
nd |
0 |
0 |
|||||||
Grade |
|||||||||
High/ grade 3 |
45 |
111 |
<0.0005 |
30 |
81 |
<0.0005 |
15 |
30 |
0.033 |
Non-high/ grade 1 or 2 |
5 |
69 |
1 |
38 |
4 |
31 |
|||
nd |
16 |
83 |
3 |
1 |
13 |
82 |
|||
Size |
|||||||||
≤20 mm |
16 |
87 |
0.054 |
12 |
65 |
0.113 |
4 |
22 |
0.392 |
>20 mm |
33 |
93 |
20 |
55 |
13 |
38 |
|||
Age (years) |
|||||||||
<50 |
9 |
54 |
0.105 |
5 |
36 |
0.120 |
4 |
18 |
0.757 |
≥50 |
39 |
119 |
28 |
85 |
11 |
34 |
|||
Lymph node status |
|||||||||
Positive |
N/A |
N/A |
N/A |
8 |
23 |
0.777 |
N/A |
N/A |
N/A |
Negative |
8 |
29 |
|||||||
ER status |
|||||||||
Positive |
24 |
142 |
0.008 |
6 |
45 |
0.038 |
18 |
97 |
0.138 |
Negative |
41 |
113 |
28 |
75 |
13 |
38 |
|||
nd |
1 |
8 |
0 |
0 |
1 |
8 |
|||
PR status |
|||||||||
Positive |
16 |
117 |
0.003 |
3 |
36 |
0.013 |
13 |
81 |
0.153 |
Negative |
48 |
138 |
31 |
83 |
17 |
55 |
|||
nd |
2 |
2 |
0 |
1 |
2 |
7 |
|||
HER2 amplification |
|||||||||
Amplified |
17 |
37 |
0.039 |
7 |
14 |
0.255 |
10 |
23 |
0.077 |
Non-amplified |
48 |
220 |
27 |
106 |
21 |
114 |
|||
nd |
1 |
6 |
0 |
0 |
1 |
6 |
|||
nd no data available, N/A non-applicable.
DISCUSSION
In vitro studies have shown important regulatory functions for LRH-1 in breast cancer cell lines [1, 7, 8, 11, 13, 14]. However, little is known about LRH-1 in clinical breast cancer samples or its role in the biology of breast cancer, its relationship to in situ to invasive transition, how this might be regulated or its association with tumour behaviour. In this study, LRH-1 expression in breast cancer was explored, including relationships between methylation and gene expression; between gene expression, HER2 and hormonal status; and between protein expression and breast cancer characteristics.
To explore the role of methylation in the regulation of the predominantly expressed NR5A2 transcript, variant 4, we identified the presence of CpG islands within the NR5A2 gene, including in the presumed promoter region and the gene body of variant 4 itself. The presence of these CpG islands suggested that DNA methylation may have an important role in regulating NR5A2 gene expression. In keeping with this theory, analysis of TCGA Infinium HumanMethylation450 array data revealed that methylation of CpG2 was associated with lower levels of NR5A2 variant 4 expression, whereas methylation of CpG3 was associated with higher NR5A2 variant 4 expression. Although this appears paradoxical, this observation is consistent with the evidence that CpG-island methylation is context-dependent, whereby methylation of CpG-island promoter sites is associated with transcriptional silencing while gene body CpG-island methylation is not associated with gene silencing and may instead be associated with transcription [24].
The expression of NR5A2 variant 4 has been reported in both ER-positive and ER-negative breast cancer cell lines [20] and our analysis of mRNA sequencing data of TCGA invasive breast cancer cohort found that variant 4 was also the predominantly expressed NR5A2 transcript in primary breast cancers. Expression of this variant was higher in ER-positive breast tumours compared with ER-negative tumours, supporting the findings of Muscat et al. [25], who in their study of 66 invasive breast cancers and 50 normal breast samples observed that NR5A2 mRNA expression was greater in ER-positive tumours compared with ER-negative tumours and was negatively correlated with tumour grade [25]. High levels of NR5A2 mRNA in ER-positive tumours may be related to the role of LRH-1 in the ER transcriptional program, but high levels of NR5A2 mRNA do not necessarily lead to high levels of LRH-1 protein or LRH-1 activity. Variant 4 is subject to E2-mediated degradation, resulting in a significantly shorter half-life in ER-positive cells compared with ER-negative cells [23]. In addition, the resultant protein has also been reported to be more stable in ER-negative breast cancer cells compared with ER-positive cells [23]. These findings support our observation of increased IHC reactivity (IHC granular pattern) in ER-negative carcinomas compared with ER-positive tumours. Furthermore, expression of NR5A2 mRNA in ER-negative tumours is strongly correlated with expression of multiple co-regulators and upregulation of ER-related genes, relationships which are not observed in ER-positive tumours [9]. Taken together, the data suggest that there is differential modulation of LRH-1 activity between ER-positive and ER-negative tumours, and that LRH-1 has roles in ER-associated, but ER-independent pathways in ER-negative tumours, similar to the observations made in breast cancer cell lines [11].
Our LRH-1 IHC data support the ER-positive/ER-negative differential. Tumours with predominantly coarse staining were more likely to be ER-negative and also display aggressive phenotypic features such as being high grade. This result is in contrast to the two previous studies examining LRH-1 expression by IHC in breast cancers that reported LRH-1 expression to be associated with favourable tumour characteristics [15, 26]. The discordant findings are likely to be due to the different IHC primary antibodies and scoring criteria used. Both the previous studies (Annicotte et al. [15] and Miki et al. [26]) used an anti-LRH-1 IHC antibody directed only towards the 541 amino acid protein resulting from the mRNA transcript NM_205860 (uc001gvb.2). This transcript was observed in TCGA breast cancer cohort to be expressed at very low levels (median 0 RSEM value, range 0-255) compared with the predominant transcript, variant 4 (median 59.9 RSEM value, range 0-1390) and therefore may not truly reflect total LRH-1 expression in breast cancers. In contrast, the antibody used in this study recognizes all forms of the LRH-1 protein. As for the scoring methodology, the IHC scoring criteria used by Annicotte et al. were not stated [15], while Miki et al. quantified only the percentage of nuclear reactivity, considering tumours with staining in at least 10% of tumour nuclei to be positive for LRH-1 expression [26].
LRH-1 is subject to various types of post-translational modifications including phosphorylation, acetylation, ubiquitination and SUMOylation [27]. SUMOylation refers to the covalent attachment of the small ubiquitin-related modifier (SUMO) to specific lysine residues [28]. SUMOylation of LRH-1 leads to sequestration of LRH-1 in promyelocytic leukaemia (PML) protein nuclear bodies, localising as discrete nuclear dots, in contrast to unSUMOylated LRH-1, which is distributed diffusely in the nucleus [29]. Sequestrated LRH-1 in PML protein nuclear bodies is transcriptionally inactive and is thought to serve as a LRH-1 reservoir [29]. Low intra-nuclear concentrations of LRH-1 lead to de-SUMOylation and release of LRH-1 from the PML protein nuclear bodies and an associated change in nuclear distribution of LRH-1 from discrete nuclear dots to a diffuse nuclear distribution [29]. Therefore the coarse granular IHC staining pattern could represent breast tumours with stored SUMOylated intra-nuclear LRH-1 protein in PML protein nuclear bodies due to high overall LRH-1 levels. The observation of coarse granular IHC staining occurring on a background of diffuse nuclear staining is consistent with this hypothesis (Figure 3).
If coarsely granular nuclear LRH-1 IHC staining is indeed indicative of tumours with higher intra-nuclear levels of LRH-1, the association of this staining pattern with aggressive phenotypic features in breast cancer is consistent with previously reported associations of LRH-1 with tumorigenic functions in breast cancer cells [1, 7, 8, 11, 14], the association of LRH-1 knockdown with down-regulation of genes that are overexpressed in high-grade tumours and associated with poor outcome [7], and with the observation that high NR5A2 mRNA expression in conjunction with low CDKN1A expression conferred poor disease-free survival in breast cancer patients in TCGA dataset [1].
In conclusion, the nuclear receptor LRH-1 has previously been reported to have tumorigenic functions in breast cancer cell lines, raising the possibility that it may have similar roles in clinical breast cancer. Analysis of TCGA cohort of invasive breast carcinomas revealed that, similar to breast cancer cell lines, variant 4 was the predominantly expressed transcript and expression of variant 4 was associated with DNA methylation status. Distinct IHC nuclear reactivity patterns were identified for LRH-1 ranging from finely dispersed to coarsely granular staining, with densely punctate and coarsely granular nuclear staining being associated with aggressive breast cancer characteristics, suggesting that LRH-1 expression is informative of breast cancer biology. This may be of clinical relevance as IHC can provide a relatively straightforward method of identifying cases potentially at higher risk of poor clinical outcome and therefore can be used to risk stratify patients for appropriate treatment selection and may be indicative of response to proposed LRH-1 antagonists [23, 30–33].
MATERIALS AND METHODS
NR5A2 mRNA expression and DNA methylation
The Cancer Genome Atlas (TCGA) breast invasive carcinoma cohort was utilized to obtain mRNA expression and DNA methylation data in a large number of breast cancer cases. Processed gene isoforms expression data (RNA-Seq version 2, level 3 data, accessed 28 January 2016), HM450 array data (level 3 data, accessed 28 January 2016), and clinical and histopathological data from invasive breast cancer cohort (breast invasive carcinoma) [34] were obtained from the TCGA data portal.
Gene isoforms expression values and methylation level of HM450 were reported as normalized RSEM (RNA-Seq by Expectation Maximization) count [35] and beta value, respectively. Quality control and batch effects analysis were performed using Relative Loge Expression plot and unsupervised method including principal component analysis (PCA).
The RNA sequencing data was examined for expression of isoforms uc009wzg.1, uc001gvb.2, uc001gvc.2, uc009wzh.2, and uc010pph.1, corresponding to the five NR5A2 transcripts, variant 4 being uc009wzh.2.
The HM450 methylation array data was examined for methylation of the CpG island associated with the presumed promoter region of variant 4 (CpG2) and of the four intragenic CpG islands (CpG3, CpG4, CpG5, CpG6), specifically methylation of two CpG2 probes (cg24352938, cg21540765), two CpG3 probes (cg04097639, cg17804356), three CpG4 probes (cg18203366, cg17520027, cg04025964), eight CpG5 probes (cg06244002, cg14025556, cg05391318, cg17486263, cg00001583, cg02773945, cg 05470502, cg22515278) and six CpG6 probes (cg00026457, cg11800251, cg01333884, cg02901753, cg17873998, cg23455785). For each probe, samples with a beta value of ≥0.2 were regarded as methylated.
TCGA cases were considered positive for ER and PR if there was reported staining in at least 10% of tumour cells. The exact percentage of positive tumour nuclei was not stated for cases with <10% reactivity, precluding the use of a 1% cut-off to define hormone receptor positivity. For HER2, cases with equivocal immunohistochemical staining were not included in the analysis.
Patient cohorts
Invasive carcinomas with associated DCIS were obtained from the Peter MacCallum Cancer Centre pathology department from archived diagnostic cases between 2000 and 2011. The DCIS cases were obtained from archived diagnostic cases from the pathology departments of Peter MacCallum Cancer Centre between 2004 and 2013, and Royal Prince Alfred Hospital between 1992 and 2004. Haematoxylin and eosin-stained sections of all cases were reviewed by a pathologist to confirm diagnosis and the tumour grade of invasive breast carcinomas was determined using modified Bloom and Richardson with grade 3 tumours considered to be high grade. DCIS cases were reviewed and nuclear grade was classified as low, intermediate, and high according to consensus guidelines [36].
LRH-1 and biomarker protein expression
LRH-1 protein expression was assessed by immunohistochemistry on 3 μm-thick sections of formalin-fixed, paraffin-embedded tissue microarrays containing DCIS and invasive breast cancer cases. Sections were de-waxed and hydrated through ethanol to water. Antigen retrieval was performed using High pH EnVision FLEX Target Retrieval Solution (Dako) at 124°C at 15-16 PSI for 4 minutes. Sections were incubated with the anti-LRH-1 rabbit polyclonal antibody (Catalog number HPA005455, Sigma Aldrich, St. Louis, MO) at 1:100 dilution overnight at 4°C, followed by detection using the EnVision FLEX/HRP DAB detection kit (Dako).
The LRH-1 antibody used is directed towards a 147 amino acid sequence present in all versions of the LRH-1 protein and its specificity was assessed by Western blot (Supplementary Figure 1). This antibody has been documented by the Human Protein Atlas database to stain epithelial cells of the majority of breast cancers, where ubiquitous nuclear staining with a speckled pattern is expected [37, 38]. ER, progesterone receptor, CK5/6, EGFR and Ki-67 immunohistochemistry and HER2 silver-enhanced in situ hybridization were performed on the LRH-1 IHC cohort using tissue microarray sections, as previously described [39, 40]. For ER and PR, tumours were regarded as positive if at least 10% of tumour nuclei were reactive, regardless of intensity. A cut-off of 10% tumour cell positivity was chosen to account for non-specific reactivity. HER2 amplification status was assessed by counting signals in twenty tumour nuclei, where possible. Tumours were considered HER2-amplified if the average number of signals per tumour nucleus was at least 6 [41].
Tumours were classified into intrinsic subtypes according to the St Gallen International Expert Consensus 2013 [42]. Luminal A-like and luminal B-like tumours were considered luminal tumours, while HER2 positive tumours without ER or PR positivity and triple negative (including basal-like) tumours were considered non-luminal tumours.
Approval for the project was obtained from the ethics committees of Peter MacCallum Cancer Centre (project numbers 02/26, 10/16, and 00/81) and Royal Prince Alfred Hospital (project HREC/11/RPAH/126).
Statistical analysis
Comparisons of continuous data between two groups was evaluated by Mann-Whitney U test. Fisher’s exact probability test was used to assess 2x2 contingency tables. For each comparison, a two-tailed p value of 0.05 or less was considered to be statistically significant. All statistical analyses were performed using IBM SPSS version 22.0 (IBM Corporation, NY).
Abbreviations
DCIS: Ductal carcinoma in situ; ER: Estrogen receptor; IHC: Immunohistochemistry; HM450: Infinium HumanMethylation450 array; LRH-1: Liver receptor homologue 1; PML: Promyelocytic leukaemia; PR: Progesterone receptor; RSEM: RNA-Seq by Expectation Maximization; TCGA: The Cancer Genome Atlas; SUMO: Small ubiquitin-related modifier.
Author contributions
J-MBP generated the IHC data, analysed the data, and wrote the manuscript. RM downloaded, processed and analysed the TCGA data, AC conceptualised the study, provided the IHC antibody, interpreted the data and critically revised the manuscript, KK conceptualised the study and interpreted the data, EAT generated the IHC data, DJB generated the IHC data, TM critically revised the manuscript, EKAM provided cases for the study and critically revised the manuscript, CSL provided cases for the study and critically revised the manuscript, SAO’T provided cases for the study and critically revised the manuscript, CC conceptualised the study and critically revised the manuscript, KLG conceptualised the study, provided TCGA data, interpreted the data and critically revised the manuscript, AD conceptualised the study, interpreted the data and critically revised the manuscript, SBF conceptualised the study, provided cases for the study, interpreted the data and critically revised the manuscript. All authors read and approved the final manuscript.
CONFLICTS OF INTEREST
The authors declare that they have no conflicts of interest.
FUNDING
National Breast Cancer Foundation (CG12-07 and CG10-04) and the Operational Infrastructure Support Program Funding of the Victorian State Government to the Olivia Newton-John Cancer Research Institute. KLG supported by the Victorian Cancer Agency.
REFERENCES
1. Bianco S, Jangal M, Garneau D, Gevry N. LRH-1 controls proliferation in breast tumor cells by regulating CDKN1A gene expression. Oncogene. 2015; 34:4509-4518.
2. Fayard E, Auwerx J, Schoonjans K. LRH-1: an orphan nuclear receptor involved in development, metabolism and steroidogenesis. Trends Cell Biol. 2004; 14:250-260.
3. Bayrer JR, Mukkamala S, Sablin EP, Webb P, Fletterick RJ. Silencing LRH-1 in colon cancer cell lines impairs proliferation and alters gene expression programs. Proc Natil Acad Sci U S A. 2015; 112:2467-2472.
4. Schoonjans K, Dubuquoy L, Mebis J, Fayard E, Wendling O, Haby C, Geboes K, Auwerx J. Liver receptor homolog 1 contributes to intestinal tumor formation through effects on cell cycle and inflammation. Proc Natl Acad Sci U S A. 2005; 102:2058-2062.
5. Fernandez-Marcos PJ, Auwerx J, Schoonjans K. Emerging actions of the nuclear receptor LRH-1 in the gut. Biochim Biophys Acta. 2011; 1812:947-955.
6. Benod C, Vinogradova MV, Jouravel N, Kim GE, Fletterick RJ, Sablin EP. Nuclear receptor liver receptor homologue 1 (LRH-1) regulates pancreatic cancer cell growth and proliferation. Proc Natl Acad Sci U S A. 2011; 108:16927-16931.
7. Bianco S, Brunelle M, Jangal M, Magnani L, Gevry N. LRH-1 governs vital transcriptional programs in endocrine-sensitive and -resistant breast cancer cells. Cancer Res. 2014; 74:2015-2025.
8. Chand AL, Herridge KA, Thompson EW, Clyne CD. The orphan nuclear receptor LRH-1 promotes breast cancer motility and invasion. Endocr Relat Cancer. 2010; 17:965-975.
9. Doan TB, Eriksson NA, Graham D, Funder JW, Simpson ER, Kuczek ES, Clyne C, Leedman PJ, Tilley WD, Fuller PJ, Muscat GE, Clarke CL. Breast cancer prognosis predicted by nuclear receptor-coregulator networks. Mol Oncol. 2014; 8:998-1013.
10. Chand AL, Herridge KA, Howard TL, Simpson ER, Clyne CD. Tissue-specific regulation of aromatase promoter II by the orphan nuclear receptor LRH-1 in breast adipose stromal fibroblasts. Steroids. 2011; 76:741-744.
11. Chand AL, Wijayakumara DD, Knower KC, Herridge KA, Howard TL, Lazarus KA, Clyne CD. The orphan nuclear receptor LRH-1 and ERalpha activate GREB1 expression to induce breast cancer cell proliferation. PLoS One. 2012; 7:e31593.
12. Zhou J, Suzuki T, Kovacic A, Saito R, Miki Y, Ishida T, Moriya T, Simpson ER, Sasano H, Clyne CD. Interactions between prostaglandin E(2), liver receptor homologue-1, and aromatase in breast cancer. Cancer Res. 2005; 65:657-663.
13. Clyne CD, Speed CJ, Zhou J, Simpson ER. Liver receptor homologue-1 (LRH-1) regulates expression of aromatase in preadipocytes. J Biol Chem. 2002; 277:20591-20597.
14. Lai CF, Flach KD, Alexi X, Fox SP, Ottaviani S, Thiruchelvam PT, Kyle FJ, Thomas RS, Launchbury R, Hua H, Callaghan HB, Carroll JS, Charles Coombes R, et al. Co-regulated gene expression by oestrogen receptor alpha and liver receptor homolog-1 is a feature of the oestrogen response in breast cancer cells. Nucleic Acids Res. 2013; 41:10228-10240.
15. Annicotte JS, Chavey C, Servant N, Teyssier J, Bardin A, Licznar A, Badia E, Pujol P, Vignon F, Maudelonde T, Lazennec G, Cavailles V, Fajas L. The nuclear receptor liver receptor homolog-1 is an estrogen receptor target gene. Oncogene. 2005; 24:8167-8175.
16. Kent WJ, Sugnet CW, Furey TS, Roskin KM, Pringle TH, Zahler AM, Haussler D. The human genome browser at UCSC. Genome Res. 2002; 12:996-1006.
17. Nitta M, Ku S, Brown C, Okamoto AY, Shan B. CPF: an orphan nuclear receptor that regulates liver-specific expression of the human cholesterol 7alpha-hydroxylase gene. Proc Natl Acad Sci U S A. 1999; 96:6660-6665.
18. Zhang CK, Lin W, Cai YN, Xu PL, Dong H, Li M, Kong YY, Fu G, Xie YH, Huang GM, Wang Y. Characterization of the genomic structure and tissue-specific promoter of the human nuclear receptor NR5A2 (hB1F) gene. Gene. 2001; 273:239-249.
19. Li M, Xie YH, Kong YY, Wu X, Zhu L, Wang Y. Cloning and characterization of a novel human hepatocyte transcription factor, hB1F, which binds and activates enhancer II of hepatitis B virus. J Biol Chem. 1998; 273:29022-29031.
20. Thiruchelvam PT, Lai CF, Hua H, Thomas RS, Hurtado A, Hudson W, Bayly AR, Kyle FJ, Periyasamy M, Photiou A, Spivey AC, Ortlund EA, Whitby RJ, et al. The liver receptor homolog-1 regulates estrogen receptor expression in breast cancer cells. Breast Cancer Res Treat. 2011; 127:385-396.
21. Pruitt KD, Brown GR, Hiatt SM, Thibaud-Nissen F, Astashyn A, Ermolaeva O, Farrell CM, Hart J, Landrum MJ, McGarvey KM, Murphy MR, O'Leary NA, Pujar S, et al. RefSeq: an update on mammalian reference sequences. Nucleic Acids Res. 2014; 42:D756-D763.
22. Gardiner-Garden M, Frommer M. CpG islands in vertebrate genomes. J Mol Biol. 1987; 196:261-282.
23. Lazarus KA, Zhao Z, Knower KC, To SQ, Chand AL, Clyne CD. Oestradiol reduces liver receptor homolog-1 mRNA transcript stability in breast cancer cell lines. Biochem Biophys Res Commun. 2013; 438:533-539.
24. Jones PA. Functions of DNA methylation: islands, start sites, gene bodies and beyond. Nat Rev Genet. 2012; 13:484-492.
25. Muscat GE, Eriksson NA, Byth K, Loi S, Graham D, Jindal S, Davis MJ, Clyne C, Funder JW, Simpson ER, Ragan MA, Kuczek E, Fuller PJ, et al. Research resource: nuclear receptors as transcriptome: discriminant and prognostic value in breast cancer. Mol Endocrinol. 2013; 27:350-365.
26. Miki Y, Clyne CD, Suzuki T, Moriya T, Shibuya R, Nakamura Y, Ishida T, Yabuki N, Kitada K, Hayashi S, Sasano H. Immunolocalization of liver receptor homologue-1 (LRH-1) in human breast carcinoma: possible regulator of insitu steroidogenesis. Cancer Lett. 2006; 244:24-33.
27. Stein S, Schoonjans K. Molecular basis for the regulation of the nuclear receptor LRH-1. Curr Opin Cell Biol. 2015; 33:26-34.
28. Yang FM, Pan CT, Tsai HM, Chiu TW, Wu ML, Hu MC. Liver receptor homolog-1 localization in the nuclear body is regulated by sumoylation and cAMP signaling in rat granulosa cells. FEBS J. 2009; 276:425-436.
29. Chalkiadaki A, Talianidis I. SUMO-dependent compartmentalization in promyelocytic leukemia protein nuclear bodies prevents the access of LRH-1 to chromatin. Mol Cell Biol. 2005; 25:5095-5105.
30. Nadolny C, Dong X. Liver receptor homolog-1 (LRH-1): a potential therapeutic target for cancer. Cancer Biol Ther. 2015; 16:997-1004.
31. Corzo CA, Mari Y, Chang MR, Khan T, Kuruvilla D, Nuhant P, Kumar N, West GM, Duckett DR, Roush WR, Griffin PR. Antiproliferation activity of a small molecule repressor of liver receptor homolog 1. Mol Pharmacol. 2015; 87:296-304.
32. Rey J, Hu H, Kyle F, Lai CF, Buluwela L, Coombes RC, Ortlund EA, Ali S, Snyder JP, Barrett AG. Discovery of a new class of liver receptor homolog-1 (LRH-1) antagonists: virtual screening, synthesis and biological evaluation. ChemMedChem. 2012; 7:1909-1914.
33. Benod C, Carlsson J, Uthayaruban R, Hwang P, Irwin JJ, Doak AK, Shoichet BK, Sablin EP, Fletterick RJ. Structure-based discovery of antagonists of nuclear receptor LRH-1. J Biol Chem. 2013; 288:19830-19844.
34. The Cancer Genome Atlas. National Institute of Health.
35. Li B, Dewey CN. RSEM: accurate transcript quantification from RNA-Seq data with or without a reference genome. BMC Bioinformatics. 2011; 12:323.
36. Consensus conference on the classification of ductal carcinoma in situ. Hum Pathol. 1997; 28:1221-1225.
37. Uhlen M, Oksvold P, Fagerberg L, Lundberg E, Jonasson K, Forsberg M, Zwahlen M, Kampf C, Wester K, Hober S, Wernerus H, Bjorling L, Ponten F. Towards a knowledge-based Human Protein Atlas. Nat Biotechnol. 2010; 28:1248-1250.
38. Uhlen M, Fagerberg L, Hallstrom BM, Lindskog C, Oksvold P, Mardinoglu A, Sivertsson A, Kampf C, Sjostedt E, Asplund A, Olsson I, Edlund K, Lundberg E, et al. Proteomics. Tissue-based map of the human proteome. Science. 2015; 347:1260419.
39. Yan M, Xu H, Waddell N, Shield-Artin K, Haviv I, McKay MJ, Fox SB. Enhanced RAD21 cohesin expression confers poor prognosis in BRCA2 and BRCAX, but not BRCA1 familial breast cancers. Breast Cancer Res. 2012; 14:R69.
40. Deb S, Jene N; Kconfab Investigators, Fox SB. Genotypic and phenotypic analysis of familial male breast cancer shows under representation of the HER2 and basal subtypes in BRCA-associated carcinomas. BMC Cancer. 2012; 12:510.
41. Wolff AC, Hammond ME, Hicks DG, Dowsett M, McShane LM, Allison KH, Allred DC, Bartlett JM, Bilous M, Fitzgibbons P, Hanna W, Jenkins RB, Mangu PB, et al. Recommendations for human epidermal growth factor receptor 2 testing in breast cancer: American Society of Clinical Oncology/College of American Pathologists clinical practice guideline update. J Clin Oncol. 2013; 31:3997-4013.
42. Goldhirsch A, Winer EP, Coates AS, Gelber RD, Piccart-Gebhart M, Thurlimann B, Senn HJ; Panel members. Personalizing the treatment of women with early breast cancer: highlights of the St Gallen International Expert Consensus on the Primary Therapy of Early Breast Cancer 2013. Ann Oncol. 2013; 24:2206-2223.