INTRODUCTION
Bladder cancer (BC) is the fourth most common cancer in men in the United States and eighth most common cause of cancer death [1]. In 2016, an estimated 76,000 men and women will be diagnosed with BC in the US and 16,000 people will die of BC [2]. Cigarette smoking (mainly exposure to aromatic amines) accounts for 50% of bladder cancers [3, 4]. Non-tobacco related occupational exposure to amines, 4-aminobiphenyl & anilines (10% of all cases), as well as phenacetin derived analgesics (oral pain medications) [5, 6] have also been known to contribute to the etiology of BC. BC originates primarily in the transitional cell epithelium (urothelial epithelium) that lines the inner surface of the bladder and is directly exposed to urine, which is also known as transitional cell carcinoma (TCC) [4]. Approximately 70-75% of newly diagnosed BCs are non-muscle invasive (NMIBC), previously referred to as “superficial” BC, while 25-30% of tumors upon initial diagnosis are muscle invasive (≥ clinical stage T2) [4]. NMIBC is typically treated with endoscopic transurethral resection (TUR), which may be followed by an intravesical therapy, depending on the extent of the cancer, tumor grade and the presence of carcinoma in situ. Given the high risk of recurrence and disease progression, careful surveillance after cancer removal by TUR via cystoscopy is currently the standard clinical practice. Intravesical therapies with Bacillus Calmette-Guerin (BCG) or chemotherapeutic agents (e.g., mitomycin C), delivered via a urethral catheter, are used to prevent or delay recurrence and progression after TUR [7]. Although BCG has been more effective than other agents, 20-40% of patients fail to respond [8]. Recurrence is common despite BCG treatment with recurrence rates for high risk T1 tumors ranging from 16 to 40% and progression rates of 4% to 40%. [9-14]. Upon diagnosis of muscle invasive bladder cancer (stage T2), the current definitive treatment is radical cystectomy (surgical extirpation of the bladder) and urinary diversion. Overall survival is poor once distant metastasis (~15% 5 year survival) has occurred [15] with stage being the most important prognostic factor of BC [16].
The clinical course for both early and late stage BC continues to be characterized by significant patient burden due to numerous occurrences and recurrences requiring frequent surveillance strategies, intravesical drug therapies, and even more aggressive treatments in patients with locally advanced or metastatic disease Additionally, BC is the most expensive overall cancer to treat given its propensity to recur and the need for frequent treatment and surveillance [17, 18]. BC thus carries a significant patient burden as well as a healthcare cost-related burden underscoring the need to optimize BC care and need for prevention strategies especially targeting non-muscle invasive patients [17, 18]. Evaluation of chemoprevention interventions in BC patients is especially feasible given physiological exposure of bladder urothelial cells to excreted compounds, readily available pathological specimens for analysis, and measurable intermediate endpoint biomarkers [17, 19]. However, other than smoking cessation, there is a paucity of research that systematically examines agents for the chemoprevention of BC [20]. Smoking cessation has been shown to decrease recurrence and improve prognosis, yet this beneficial effect is only observed for long term smoking cessation ( > 10 years) [17, 19].
The objective is to review the available evidence from epidemiological, in vitro, pre-clinical animal and early clinical trials of various agents evaluated for bladder cancer chemoprevention with a focus on sulforaphane for bladder cancer chemoprevention.
RESULTS
Current strategies for bladder cancer chemoprevention
Vitamins
Previous studies have focused on vitamin intake including, vitamin C, vitamin B6 and vitamin E and an essential trace element, selenium. However, the majority of these studies failed to indicate a promising agent for primary or secondary chemoprevention of BC [21]. Retinoids (vitamin A derivatives) and alpha tocopherol (vitamin E) have been studied as a putative chemopreventive agent in bladder cancer. In vitro studies suggested that rats with vitamin A deficiency were more likely to develop environmentally induced bladder cancer, and that supplementation of vitamin A could prevent bladder cancer development [22-25]. However, clinical studies do not support a chemopreventive role of retinoids, including the ATBC study that targeted at-risk smokers and assigned patients to beta-carotene, alpha tocopherol, both or placebo and showed no benefit in prevention of bladder cancer at 6 years of follow up [26]. A secondary analysis of the SELECT trial also failed to show a protective effect for vitamin E or selenium for bladder cancer [27]. Other studies exploring the role of retinoids for secondary chemoprevention also showed no benefit, and as a result of concerns for toxicity (increased myocardial infarction risk) and lack of clear benefit, one study was terminated prior to accrual [28-30].
Pyridoxine (B6) has been investigated in two randomized trials for secondary chemoprevention without evidence of a benefit [31, 32]. Ascorbic acid (vitamin C) has not been studied in randomized trials, and epidemiological data is not convincing with respect to its protective effect [33]. Mega dose multivitamins have not demonstrated clinical effectiveness in chemoprevention despite epidemiological research and clinical research suggesting a possible role for chemoprevention [34].
NSAIDS and Cox-2 inhibitors
More recent chemopreventive efforts have exposed the role of non-steroidal anti-inflammatory drugs, specifically the role of selective COX-2 inhibitors. This has included clinical studies with celecoxib that suggested a correlation between COX-2 expression and prognosis. One trial in nonmuscle invasive bladder cancer patients showed similar risk in progression and recurrence between celecoxib and placebo [35]. The results of an expanded Phase III clinical trial in non-muscle invasive bladder cancer patients who responded to BCG treated with celecoxib or placebo are still not available. Intriguingly, an in vitro study using allyl isothiocyanate (AITC) with celecoxib produced depletion of prostaglandin E2, a key downstream signaling molecule of Cox-2, caspase activation and down regulation of vascular endothelial growth factor in the tumor tissues. These data show that AITC and celecoxib may complement each other in inhibiting bladder carcinogenesis, providing a novel combination approach for future validation in preclinical models for chemoprevention of bladder cancer [36].
EGFR inhibitors and mTOR inhibitors
Also, research is being conducted with erlotinib, highly selective, reversible inhibitor of epidermal growth factor receptor (HER1/EGFR) tyrosine kinase which is overexpressed in more than 75% of bladder cancers [37]. One phase 2 clinical trial involved neoadjuvant administration of erlotinib in patients before undergoing radical cystectomy with a complete response rate in twenty five percent of patients. There was substantial skin toxicity noted especially in patients who experienced complete response [38]. A phase IIa randomized multi-institutional trial (NCT02169284) is ongoing investigating role of erlotinib in presurgical (RC or TURBT) patients [39] .
Additionally, research has focused on mTOR inhibition as a potential target for chemoprevention in bladder cancer [40]. Metformin, a commonly utilized diabetes medication has properties as an mTOR inhibitor and has been investigated with underwhelming results mostly in a retrospective fashion [41, 42]. One nonrandomized clinical trial comparing non muscle invasive BC patients taking metformin to placebo showed no difference in recurrence and no statistical difference in time to recurrence [43]. Also, in vitro research using Rhodiola rosea extract has shown to inhibit mTOR and decrease growth of bladder cancer [42]. Finally, some research has demonstrated a possible role of estrogen receptor blockade using tamoxifen in mouse model in modulating bladder tumorigenesis [44].
Soy isoflavones
Isoflavones, which are soy derivatives, have recently been investigated as a possible chemoprevention agent based on epidemiological and in vitro evidence. A recent trial by Messing et al., explored the use of genistein, an isoflavone, in pre-surgical bladder cancer patients and demonstrated significant inhibition of p-EGFR at dose-specific levels, but other apoptotic and proliferative biomarkers were not impacted [45]. There has been an increase in newer research that has focused on isolation of (natural) bioactive compounds. Recently, Justicidin A, a methanol extract of Justicia procumbens, has been investigated in vitro as an anti-angiogenic and apoptosis-inducing agent [46], as have pomegranate extracts [47]. Additionally, green tea catechins have been demonstrated in several in vitro and in vivo studies to have significant anti-carcinogenic potential [48-51]. However, these are early observations that have to be further validated in preclinical models prior to evaluation in clinical trials.
Despite attempts at identifying other single and combination agents for chemoprevention, several epidemiological, in vitro, preclinical, and early phase trials have shown that the phytochemicals, isothiocyanates (ITCs), specifically sulforaphane (SFN) present in Brassicaceae or “cruciferous” vegetables in the precursor form of glucoraphanin [52-54], are: (a) available in standardized formulations; (b) bioavailable - both systemically and in the bladder; (c) observed to be potent inhibitors of BC carcinogenesis through multiple mechanisms [2]; and (d) associated with no dose-limiting toxicities at the proposed dose levels, thus supporting further development of SFN in phase I/II human studies targeting bladder cancer.
Sulforaphane for bladder cancer chemoprevention
Sulforaphane (SFN), (-)-1-isothiocyanato-(4R)-(methylsulfinyl) butane [CH3-SO-(CH2)4-NCS], is an isothiocyanate found in high concentrations in broccoli sprouts. Sulforaphane was first isolated and shown to be a potent anti-carcinogenic agent in 1992 by Zhang, et al. [54].
Epidemiological evidence for sulforaphane for bladder cancer chemoprevention
Epidemiological studies have shown a potential role for increased fluid intake and consumption of cruciferous vegetables, particularly for broccoli consumption, in reducing the risk of BC [55-57]. In a large prospective study, 39% reduction in BC risk was observed with an intake of 2 servings or more of broccoli compared to < 1 serving per week (p = 0.0009) [55]. In a meta-analysis of ten clinical trials, cruciferous vegetable intake was associated with decreased risk of bladder cancer overall [58].
Pharmacokinetics of sulforaphane
Given the epidemiological studies which suggested a potential role for SFN as a chemoprevention agent, many studies have been conducted to elicit the pharmacokinetics of SFN. Broccoli accumulates significant amounts of the phytonutrient glucoraphanin (4-methylsulfinylbutyl glucosinolates), which is metabolized in vivo to the biologically active sulforaphane. This conversion requires myrosinase, which is present in the plant as well as in the gastrointestinal tract [53]. Upon being consumed, SFN is metabolized via the mercapturic acid pathway to form cysteinylglycine-, cysteine-, and N-acetylcysteine (NAC) conjugates. These metabolites are then excreted via the urine [59, 60]. Studies have shown that 70% of an initial SFN dose was able to be retrieved in urine [61, 62]. Urine has been shown in rat models to have significant concentration of SFN present as NAC conjugates with 72% to 95% of the original SFN dose recovered in urine [63] Also, bladder tissue in a rat model has been shown to have very high concentrations of SFN after gastric lavage, second in visceral organ concentration only to stomach tissue [64]. Uptake of SFN into bladder cancer cells is dependent upon diffusion and rapidly conjugate with GSH and other intracellular proteins [65]. Several factors including concentration, lipophilicity, and exposure time influence uptake [65-67].
Preclinical studies using animal models have demonstrated bioavailability of SFN with metabolites distributed to all tissues, including the bladder, suggesting the potential for systemic benefits [68-70]. Administration of a freeze-dried aqueous extract of broccoli sprouts to rats significantly and dose-dependently inhibited bladder cancer development induced by N-butyl-N-(4-hydroxybutyl) nitrosamine [71]. The incidence, multiplicity, size, and progression of BC were all inhibited by the extract, while the extract itself caused no histologic changes in the bladder. Moreover, inhibition of bladder carcinogenesis by the extract was associated with significant induction of phase-II enzymes such as glutathione S-transferase and NAD(P)H:quinone oxidoreductase 1 in the bladder. Over 70% of the isothiocyanates present in the extract were excreted in the urine as isothiocyanate equivalents (isothiocyanates + dithiocarbamates) within 12 h after a single oral dose, indicating high bioavailability and rapid urinary excretion. Urinary concentrations in extract-treated rats were 2 to 3 orders of magnitude higher than those in plasma, indicating that the bladder epithelium, the major site of bladder cancer development, is most exposed to orally-dosed isothiocyanate. In a murine UMUC3 xenograft model, semi-purified diets containing 4% broccoli sprouts, or 2% broccoli sprout isothiocyanate extract, or gavaged pure SFN, or erucin (each at 295 μmol/kg, similar to dietary exposure) produced tumor weight reduction of 42% (p = 0.02), 42% (p = 0.04), 33% (p = 0.04), and 58% (p < 0.0001), respectively. SFN and erucin metabolites are present in mouse plasma (micromolar range) and tumor tissue, with N-acetylcysteine conjugates as the most abundant [68].
Several clinical trials have been conducted to evaluate the effectiveness of SFN for chemoprevention, most of which have investigated bioavailability in healthy, disease-free subjects [72-75,125]. The method of ingestion in these clinical trials has varied from pure SFN, broccoli soups/pill forms, and broccoli as a food item. Glucoraphanin (GRR) in broccoli is converted to SFN either by plant myrosinases, or if the plant myrosinases have been denatured by cooking, by bacterial myrosinases in the human colon. SFN is passively absorbed and rapidly conjugated with glutathione by glutathione S-transferases (GSTs), then metabolized sequentially by γ-glutamyl-transpeptidase (GTP), cysteinyl-glycinease (GCase) and N-acetyltransferase (NAT). The conjugates are actively transported into the systemic circulation where the mercapturic acid and its precursors are urinary excretion products. Deconjugation may also occur to yield the parent isothiocyanate, SFN. The mercapturic acid and cysteine conjugate forms are the major urinary metabolites of SFN [76]. In interventions with glucosinolate-containing Brussels sprouts for 1-3 weeks, increased GST enzyme activity with increased GST-alpha induction was observed in plasma and tissues including liver, bladder, and small intestine [72, 77]. Bioavailability, as measured by urinary excretion of SFN and its metabolites (in approximately 12-hour collections after dosing), was substantially greater with the SFN-rich (mean = 70%) than with GRR-rich (mean = 5%) beverages. Inter-individual variability in excretion was considerably lower with SFR than with GRR beverage [62]. These studies have also corroborated the critical role of myrosinase in metabolizing SFN, as patients taking food sources vs. extract of SFN without myrosinase had a fourfold increase in urinary concentration [76].
Table 1: Clinical trials assessing sulforaphanes for cancer chemoprevention
Agent |
Dose/Duration |
Cancer |
Sample size |
Endpoints |
60 mg (340 μmol) “stabilized SFN” (Prostaphane®) vs. Placebo [73] |
6 months (RCT) |
Prostate (Rising PSA after prostatectomy) |
N=81 |
Lower Log PSA slope in the SFN group (p = 0.01)and serum PSA (p=<0.05) compared to placebo |
200 μmol daily [105] |
5 months |
Prostate (Rising PSA after prostatectomy) |
N=20 |
1 of 20 patients with 50% decline in PSA at 5 months |
400 g broccoli/week vs. 400 g peas/week [123] |
12 months |
Prostate (Patients with high-grade prostate intraepithelial neoplasia) (HGPIN), |
N=22 |
Significant changes in TGFβ, Insulin signaling and EGF receptor pathways |
Glucoraphanin (30 mg GFN BroccoMax™ vs. placebo)[76] |
2-8 weeks |
Breast (Abnormal mammograms/pre-biopsy) |
N=27 |
Ki-67 (p = 0.003) and HDAC3 (p = 0.044) levels significantly decreased in benign tissue. GFN supplementation was associated with a significant decrease in PBMC HDAC activity (p = 0.04). |
Pharmacodynamics/ mechanisms of action of sulforaphane
Although several molecular targets in cellular and animal models have been identified, the most sensitive target for SFN is Keap1, a key sensor for the adaptive stress response system regulated through the transcription factor Nrf2 (nuclear factor (erythroid-derived2)-like 2). Keap1 is a sulfhydryl-rich protein that represses Nrf2 signaling by facilitating the polyubiquitination of Nrf2, thereby enabling its subsequent proteasomal degradation. Interaction of SFN with Keap1 disrupts this function and allows for nuclear accumulation of Nrf2 and activation of its transcriptional program (Figure 1). Enhanced transcription of Nrf2 target genes provokes a strong cytoprotective response that enhances resistance to carcinogenesis and other diseases mediated by exposures to electrophiles and oxidants [78, 79,109]. Inhibition of Phase I enzymes by SFN may contribute to inhibition of procarcinogens and its chemopreventitive effect. The Nrf2 transcription factor is essential for induction of phase 2 proteins [78, 79, 109]. In in vitro and in vivo using murine models in oral carcinogenesis, SFN was demonstrated to activate the NRF2 pathway and downregulate oxidative damage [110-11]. SFN has also been observed to be a potent inducer of phase 2 detoxification enzymes [e.g., glutathione transferases, epoxide hydrolase, NAD(P)H: quinone reductase, and glucuronosyltransferases], which may offer protection against carcinogenesis, mutagenesis, and other forms of toxicity elicited by electrophiles and reactive forms of oxygen [69, 79-82] . SFN promotes reactive oxygen species formation by inhibition of Complex III within mitochondria [112] . Evidence suggests that SFN activation of production of 4-hydroxynonenal (a lipid peroxidation product) may be essential in the chemopreventitive mechanism. 4-hydroxynonenal induces proteins critical to apoptosis, promotes cell cycle arrest, and also activates Nrf2 pathway [113]. Other mechanisms include down regulation of NFkB, resulting in induction of cell cycle arrest and apoptosis [83], while selectively targeting abnormal/malignant cells [70] compared to normal BC cells [2, 36] . Using bladder cancer cell lines, Abbaoui, et al. [68] demonstrated that downregulation of survivin, epidermal growth factor receptor (EGFR), and human epidermal growth factor receptor 2 (HER2/neu) induced G2/M cell cycle accumulation and apoptosis [68]. In osteosarcoma cell lines, SFN decreased cell invasion and also focal adhesion kinase (FAK), both of which are important in cancer progression [114]. SFN has been shown to inhibit inflammatory responses, including downregulation of cyclooxygenase-2 (Cox-2) and reduction of prostaglandin E2 levels [84, 85]. SFN likely effects immune and inflammatory response specifically via Toll-like receptor (TLR4) suppression via blockade of thiol-mediated conjugation in macrophages [86]. Telomere length which is essential for cell survival may be decreased by exposure to SFN [115].
Other studies suggest a key mechanism involving SFN is disruption of mitochondria. SFN has been shown to promote translocation of Bak, leading to loss of transmembrane potential, mitochondrial degradation, and release of cytochrome C [78, 87]. Studies using bladder cancer cell lines suggest an M phase arrest as the primary mechanism for SFN [88]. In cervical cancer cell lines, SFN was shown to cause G2/M arrest via cyclinB1 downregulation [116]. Therefore, SFN acts upon a multitude of growth-regulatory and inflammatory pathways, some of which contain oncogenic targets, to exert its potent anti-cancer effects.
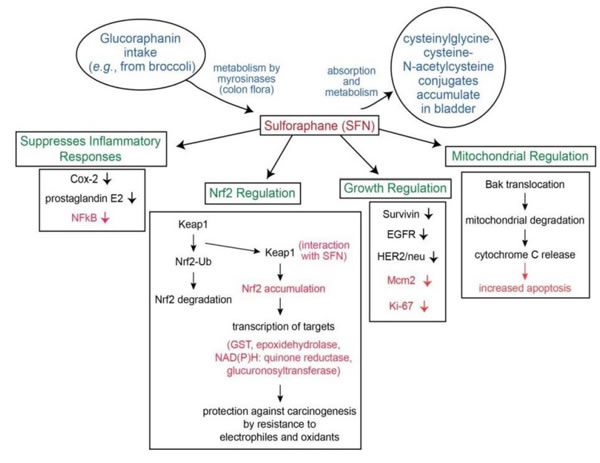
Figure 1: Metabolism and mechanism of SFN for bladder cancer chemoprevention
In vitro models in bladder cancer
In vitro studies in BC [2, 89, 90], lung [91, 121], prostate [92, 93], colorectal, thyroid [124], renal [113], ovarian [118] and leukemia cells lines [94, 95] have shown SFN to be a potent inhibitor of carcinogenesis through several molecular mechanisms [79], including in BC as discussed below. SFN inhibits the survival and proliferation of a wide array of animal and human BC cell lines [2, 96-98]. Also, SFN has been shown to be selectively more toxic to malignant urothelial cells (human) than normal urothelium [68]. Using BIU87 bladder cell lines Dang et al demonstrated that SFN downregulated NF-KB levels and upregulated Insulin-like growth factor-binding protein-3 (IGFBP-3) resulting in more apoptosis [99]. Islam et al, studied combination of SFN with acetazolamide in bladder cancer in vitro cell lines and xenografts. The authors showed diminished Ki-67, PHH3, and cyclin D1 and increases in cell cycle inhibitors p21 and p27 as well as decreases in Akt kinase activity [117]. Additionally, the studies noted downregulation of key metastatic proteins such as E-cadherin, N-cadherin, and vimentin.
Animal models
Other preclinical trials have demonstrated positive results using murine models with SFN for breast [100], skin [101], prostate [92] , oral [110], and pancreatic [70] cancers. Three studies have been conduct in mice and rats investigating the effect of SFN using a BC model. Abbaoui et al, using a mouse xenograft model showed G2/M phase arrest in models treated SFN [68]. Wang et al using the same murine xenograft documented tumor growth inhibition by 63% as well as a reduction in angiogenesis and an increase in immunological cellular response [103]. Using a rat orthotopic model, Munday et al demonstrated reduced incidence of BC by 61% and a decrease in tumor size with a delay in time to tumor progression [71].
Animal studies have suggested potential toxicity of isothiocyanates, including bladder hyperplasia [104]. However, this effect was likely secondary to overdosing. In fact, SFN have never been implicated as toxic to the bladder even in animal models using doses significantly higher than those used in clinical trials.
Clinical trials in other cancer models
To date, data from early clinical trials with SFN have focused on prostate cancer chemoprevention [73, 105, 123] with the exception of a single trial in women with abnormal mammograms, using a pre-biopsy window of opportunity for intervention with SFN [120]. The doses and formulations of SFN and duration of intervention, target populations and biomarkers/endpoints selected in these trials vary significantly. In one study, 60 mg (340 μmol) “stabilized SFN” (Prostaphane®) vs. placebo [73] was used targeting men with biochemical recurrence of prostate cancer. A reduction in serum PSA was observed in 8/20 (40%) of prostate cancer patients with no clinical toxicities, compared to placebo [105].
Targeting a similar population with 200 μmol daily for 5 months , Alumkal et al [105]demonstrated that 1 of 20 patients had a 50% decline in serum PSA at 5 months.
Using 400 g broccoli/week vs. 400 g peas/week targeting men with high-grade prostate intraepithelial neoplasia) (HGPIN), Traka et al [123] showed significant changes in TGFβ, Insulin signaling and EGF receptor pathways.
A recent double blinded randomized trial in women scheduled for breast cancer patients compared glucoraphanin supplement (Glucoraphanin (30 mg GFN BroccoMax™ vs. placebo) [76] prior to women undergoing breast biopsy. The trial showed a significant difference in several relevant biomarkers including Ki-67 and HDAC3 [120]. To our knowledge, there are no clinical trials that have been reported using SFN for BC chemoprevention.
DISCUSSION
There are some limitations that are inherent to using agents that target multiple mechanistic pathways that contribute chemoprevention effects. A pragmatic future direction to chemoprevention is to utilize a broad spectrum approach [106] that involves using bioactive formulations of botanicals (as single agents or in combination with other botanicals or biologics) that have been shown to produce robust targeting of relevant and multiple, but well characterized molecular pathways, without clinically limiting toxicity - an approach that may be more effective than agents evaluated to date. SFN, similar to other botanicals, target multiple signal transduction pathways that make it challenging to determine exactly the interplay of these mechanisms in chemoprevention. Future studies should continue to examine specifc mechanisms as well as the interplay of these mechanisms involved in modulating bladder carcinogenesis. Further characterization of the chemopreventive properties of sulforaphane is critical to our understanding these mechanisms that may identify targeted pathways of sulforaphane in bladder carcinogenesis. In addition, these findings may identify and inform selection of biomarkers in evaluating efficacy of sulforaphane in modulating bladder carcinogenesis.
Additionally, bladder cancer represents a very genetically heterogeneous cancer with more than fifteen oncogenes identified and more than 20 tumor suppressor genes with varying frequencies [107], As such, chemopreventive agents that are effective in some types of bladder cancers, may not be as effective in others. It is also recognized that these agents may have specific effects at different stages of tumor progression. This certainly seems to be true of NRF2 activation in many types of cancer including lung cancer, where advanced disease is actually associated with the accumulation of genetic alterations in KEAP1 and NF2EL2 that promote constitutive activation. (TCGA also identified similar NF2EL2 mutations in muscle-invasive bladder cancers) [121]. Thus, data obtained from any agent, including SFN, which is evaluated for a specific stage of tumor, may not be generalizable for all stages of tumor [108]. Future studies should include early phase trials including heterogenous patient populations, including non-muscle invasive and muscle-invasive BC patients, potentially block randomized to examine effectiveness in various stages of bladder tumorogenesis, Additionally, the effectiveness as well as the biological effects of these early trials should include bladder tumor cells as well as the normal appearing urothelium adjacent to bladder neoplasia since these data may inform strategies for preventing the field cancerization effect seen in progressive, high grade superficial bladder cancer. Thus, by using a well rationalized, systematic approach, rigorous experimental design that addresses challenges and limitations are critical to evaluating agents for cancer chemoprevention in early phase trials, prior to advancing to phase III trial.
To date, data from early clinical trials with SFN have focused on prostate cancer chemoprevention [73, 105, 123] with the exception of a single trial in women with abnormal mammograms, using a pre-biopsy window of opportunity for intervention with SFN [120]. The doses and formulations of SFN and duration of intervention, target populations and biomarkers/endpoints selected in these trials vary significantly. Other than smoking cessation, there is a paucity of research that systematically examines specific agents relevant for chemoprevention of bladder cancers. Despite extensive knowledge of potential targets of SFN (Nrf2 induction and downregulation of NF-kB), and resulting modulation of intermediate endpoint biomarkers (apoptosis and proliferation) relevant to bladder carcinogenesis, these agents have not been evaluated in clinical trials for bladder cancer chemoprevention. Future clinical trials should be informed from the promising pharmacokinetics and pharmacodynamics evidence that exists from current in vitro and preclinical studies to inform design of early phase chemoprevention trials target men and women with non-muscle invasive as well as invasive bladder cancers.
The clinical course for both early and late stage BC continues to be characterized by significant patient burden due to numerous occurrences and recurrences requiring frequent surveillance strategies, intravesical drug therapies, and even more aggressive treatments in patients with locally advanced or metastatic disease. Based on available evidence from epidemiological, in vitro, preclinical, and early phase trials, phytochemicals, specifically isothiocyanates (ITCs) such as sulforaphane (SFN) represent a promising potential chemopreventative agent in bladder cancer. These studies will ultimately inform development of chemoprevention interventions in both early (non-muscle invasive) and late invasive stage BC patients.
materials and METHODS
MEDLINE (Ovid), EMBASE (Ovid), AMED (Ovid), CINAHL (EBSCO) and the Cochrane Library databases were searched for epidemiological studies, in vitro, in vivo and phase I-II clinical trials on the topic of bladder cancer chemoprevention with a focus on sulforaphane in bladder cancer chemoprevention. Only clinical trials investigating sulforaphane for cancer chemoprevention that were completed and results published in the identified databases were located.
Conflicts of Interest
The authors of this manuscript have no potential conflicts of interest to report.
GRANT SUPPORT
Funding for this work is supported by the National Institute of Health Cancer Center Support Grant P30 CA076292-17.
REFERENCES
1. Siegel RL, Miller KD, Jemal A. Cancer statistics, 2016. CA Cancer J Clin. 2016; 66:7-30.
2. Veeranki OL, Bhattacharya A, Tang L, Marshall JR, Zhang Y. Cruciferous vegetables, isothiocyanates, and prevention of bladder cancer. Curr Pharmacol Rep. 2015; 1:272-282.
3. Smith ZL, Christodouleas JP, Keefe SM, Malkowicz SB, Guzzo TJ. Bladder preservation in the treatment of muscle-invasive bladder cancer (MIBC): a review of the literature and a practical approach to therapy. BJU Int. 2013; 112:13-25.
4. Witjes JA, Comperat E, Cowan NC, De Santis M, Gakis G, Lebret T, Ribal MJ, Van der Heijden AG, Sherif A, European Association of U. EAU guidelines on muscle-invasive and metastatic bladder cancer: summary of the 2013 guidelines. Eur Urol. 2014; 65:778-792.
5. Burger M, Catto JW, Dalbagni G, Grossman HB, Herr H, Karakiewicz P, Kassouf W, Kiemeney LA, La Vecchia C, Shariat S, Lotan Y. Epidemiology and risk factors of urothelial bladder cancer. Eur Urol. 2013; 63:234-241.
6. Freedman ND, Silverman DT, Hollenbeck AR, Schatzkin A, Abnet CC. Association between smoking and risk of bladder cancer among men and women. JAMA. 2011; 306:737-745.
7. Clark PE, Agarwal N, Biagioli MC, Eisenberger MA, Greenberg RE, Herr HW, Inman BA, Kuban DA, Kuzel TM, Lele SM, Michalski J, Pagliaro LC, Pal SK, et al. Bladder cancer. J Natl Compr Canc Netw. 2013; 11:446-475.
8. Hilton WM, Ercole B, Parekh DJ, Sonpavde G, Ghosh R, Svatek RS. Efficacy of combined intravesical immunotherapy and chemotherapy for non-muscle invasive bladder cancer. Expert Rev Anticancer Ther. 2011; 11:949-957.
9. Cookson MS, Sarosdy MF. Management of stage T1 superficial bladder cancer with intravesical bacillus Calmette-Guerin therapy. J Urol. 1992; 148:797-801.
10. Gohji K, Nomi M, Okamoto M, Takenaka A, Hara I, Okada H, Arakawa S, Fujii A, Kamidono S. Conservative therapy for stage T1b, grade 3 transitional cell carcinoma of the bladder. Urology. 1999; 53:308-313.
11. Herr HW. Tumour progression and survival in patients with T1G3 bladder tumours: 15-year outcome. Br J Urol. 1997; 80:762-765.
12. Hurle R, Losa A, Manzetti A, Lembo A. Intravesical bacille Calmette-Guerin in Stage T1 grade 3 bladder cancer therapy: a 7-year follow-up. Urology. 1999; 54:258-263.
13. Jimenez-Cruz JF, Vera-Donoso CD, Leiva O, Pamplona M, Rioja-Sanz LA, Martinez-Lasierra M, Flores N, Unda M. Intravesical immunoprophylaxis in recurrent superficial bladder cancer (Stage T1): multicenter trial comparing bacille Calmette-Guerin and interferon-alpha. Urology. 1997; 50:529-535.
14. Pansadoro V, Emiliozzi P, Defidio L, Donadio D, Florio A, Maurelli S, Lauretti S, Sternberg CN. Bacillus Calmette-Guerin in the treatment of stage T1 grade 3 transitional cell carcinoma of the bladder: long-term results. J Urol. 1995; 154:2054-2058.
15. von der Maase H, Sengelov L, Roberts JT, Ricci S, Dogliotti L, Oliver T, Moore MJ, Zimmermann A, Arning M. Long-term survival results of a randomized trial comparing gemcitabine plus cisplatin, with methotrexate, vinblastine, doxorubicin, plus cisplatin in patients with bladder cancer. J Clin Oncol. 2005; 23:4602-4608.
16. Berger K. [Prevention of fatal complications after smallpox vaccination. Successful measures in Austria (1951-1969)]. [Article in German]. Munch Med Wochenschr. 1970; 112:1946-1952.
17. Jacobs BL, Lee CT, Montie JE. Bladder cancer in 2010: how far have we come? CA Cancer J Clin. 2010; 60:244-272.
18. Svatek RS, Hollenbeck BK, Holmang S, Lee R, Kim SP, Stenzl A, Lotan Y. The economics of bladder cancer: costs and considerations of caring for this disease. Eur Urol. 2014; 66:253-262.
19. Rink M, Furberg H, Zabor EC, Xylinas E, Babjuk M, Pycha A, Lotan Y, Karakiewicz PI, Novara G, Robinson BD, Montorsi F, Chun FK, Scherr DS, et al. Impact of smoking and smoking cessation on oncologic outcomes in primary non-muscle-invasive bladder cancer. Eur Urol. 2013; 63:724-732.
20. Lattouf JB. Chemoprevention in bladder cancer: What’s new? Can Urol Assoc J. 2009; 3:S184-187.
21. Golijanin DJ, Kakiashvili D, Madeb RR, Messing EM, Lerner SP. Chemoprevention of bladder cancer. World J Urol. 2006; 24:445-472.
22. Becci PJ, Thompson HJ, Grubbs CJ, Squire RA, Brown CC, Sporn MB, Moon RC. Inhibitory effect of 13-cis-retinoic acid on urinary bladder carcinogenesis induced in C57BL/6 mice by N-butyl-N-(4-hydroxybutyl)-nitrosamine. Cancer Res. 1978; 38:4463-4466.
23. Cohen SM, Wittenberg JF, Bryan GT. Effect of avitaminosis A and hypervitaminosis A on urinary bladder carcinogenicity of N-(4-(5-Nitro-2-furyl)-2-thiazolyl)formamide. Cancer Res. 1976; 36:2334-2339.
24. Mathews-Roth MM, Lausen N, Drouin G, Richter A, Krinsky NI. Effects of carotenoid administration on bladder cancer prevention. Oncology. 1991; 48:177-179.
25. Sporn MB, Squire RA, Brown CC, Smith JM, Wenk ML, Springer S. 13-cis-retinoic acid: inhibition of bladder carcinogenesis in the rat. Science. 1977; 195:487-489.
26. Virtamo J, Edwards BK, Virtanen M, Taylor PR, Malila N, Albanes D, Huttunen JK, Hartman AM, Hietanen P, Maenpaa H, Koss L, Nordling S, Heinonen OP. Effects of supplemental alpha-tocopherol and beta-carotene on urinary tract cancer: incidence and mortality in a controlled trial (Finland). Cancer Causes Control. 2000; 11:933-939.
27. Lotan Y, Goodman PJ, Youssef RF, Svatek RS, Shariat SF, Tangen CM, Thompson IM JR, Klein EA. Evaluation of vitamin E and selenium supplementation for the prevention of bladder cancer in SWOG coordinated SELECT. J Urol. 2012; 187:2005-2010.
28. Alfthan O, Tarkkanen J, Grohn P, Heinonen E, Pyrhonen S, Saila K. Tigason (etretinate) in prevention of recurrence of superficial bladder tumors. A double-blind clinical trial. Eur Urol. 1983; 9:6-9.
29. Decensi A, Torrisi R, Bruno S, Costantini M, Curotto A, Nicolo G, Malcangi B, Baglietto L, Bruttini GP, Gatteschi B, Rondanina G, Varaldo M, Perloff M, et al. Randomized trial of fenretinide in superficial bladder cancer using DNA flow cytometry as an intermediate end point. Cancer Epidemiol Biomarkers Prev. 2000; 9:1071-1078.
30. Studer UE, Biedermann C, Chollet D, Karrer P, Kraft R, Toggenburg H, Vonbank F. Prevention of recurrent superficial bladder tumors by oral etretinate: preliminary results of a randomized, double blind multicenter trial in Switzerland. J Urol. 1984; 131:47-49.
31. Byar D, Blackard C. Comparisons of placebo, pyridoxine, and topical thiotepa in preventing recurrence of stage I bladder cancer. Urology. 1977; 10:556-561.
32. Newling DW, Robinson MR, Smith PH, Byar D, Lockwood R, Stevens I, De Pauw M, Sylvester R. Tryptophan metabolites, pyridoxine (vitamin B6) and their influence on the recurrence rate of superficial bladder cancer. Results of a prospective, randomised phase III study performed by the EORTC GU Group. EORTC Genito-Urinary Tract Cancer Cooperative Group. Eur Urol. 1995; 27:110-116.
33. Jacobs EJ, Henion AK, Briggs PJ, Connell CJ, McCullough ML, Jonas CR, Rodriguez C, Calle EE, Thun MJ. Vitamin C and vitamin E supplement use and bladder cancer mortality in a large cohort of US men and women. Am J Epidemiol. 2002; 156:1002-1010.
34. Nepple KG, Lightfoot AJ, Rosevear HM, O’Donnell MA, Lamm DL, Bladder Cancer Genitourinary Oncology Study G. Bacillus Calmette-Guerin with or without interferon alpha-2b and megadose versus recommended daily allowance vitamins during induction and maintenance intravesical treatment of nonmuscle invasive bladder cancer. J Urol. 2010; 184:1915-1919.
35. Sabichi AL, Lee JJ, Grossman HB, Liu S, Richmond E, Czerniak BA, De la Cerda J, Eagle C, Viner JL, Palmer JL, Lerner SP. A randomized controlled trial of celecoxib to prevent recurrence of nonmuscle-invasive bladder cancer. Cancer Prev Res (Phila). 2011; 4:1580-1589.
36. Bhattacharya A, Li Y, Shi Y, Zhang Y. Enhanced inhibition of urinary bladder cancer growth and muscle invasion by allyl isothiocyanate and celecoxib in combination. Carcinogenesis. 2013; 34:2593-2599.
37. Chow NH, Chan SH, Tzai TS, Ho CL, Liu HS. Expression profiles of ErbB family receptors and prognosis in primary transitional cell carcinoma of the urinary bladder. Clin Cancer Res. 2001; 7:1957-1962.
38. Pruthi RS, Nielsen M, Heathcote S, Wallen EM, Rathmell WK, Godley P, Whang Y, Fielding J, Schultz H, Grigson G, Smith A, Kim W. A phase II trial of neoadjuvant erlotinib in patients with muscle-invasive bladder cancer undergoing radical cystectomy: clinical and pathological results. BJU Int. 2010; 106:349-354.
39. https://clinicaltrials.gov/ct2/show/NCT02169284?term = NCT02169284&rank = 1.
40. Makhlin I, Zhang J, Long CJ, Devarajan K, Zhou Y, Klein-Szanto AJ, Huang M, Chernoff J, Boorjian SA. The mTOR pathway affects proliferation and chemosensitivity of urothelial carcinoma cells and is upregulated in a subset of human bladder cancers. BJU Int. 2011; 108:E84-90.
41. Mamtani R, Pfanzelter N, Haynes K, Finkelman BS, Wang X, Keefe SM, Haas NB, Vaughn DJ, Lewis JD. Incidence of bladder cancer in patients with type 2 diabetes treated with metformin or sulfonylureas. Diabetes Care. 2014; 37:1910-1917.
42. Liu Z, Li X, Simoneau AR, Jafari M, Zi X. Rhodiola rosea extracts and salidroside decrease the growth of bladder cancer cell lines via inhibition of the mTOR pathway and induction of autophagy. Mol Carcinog. 2012; 51:257-267.
43. Heidari F, Abbas Zade S, Mir Hosseini SH, Ghadian A. Metformin for the Prevention of Bladder Cancer Recurrence: Is it Effective? Nephrourol Mon. 2016; 8:e30261.
44. George SK, Tovar-Sepulveda V, Shen SS, Jian W, Zhang Y, Hilsenbeck SG, Lerner SP, Smith CL. Chemoprevention of BBN-Induced Bladder Carcinogenesis by the Selective Estrogen Receptor Modulator Tamoxifen. Transl Oncol. 2013; 6:244-255.
45. Messing E, Gee JR, Saltzstein DR, Kim K, diSant’Agnese A, Kolesar J, Harris L, Faerber A, Havighurst T, Young JM, Efros M, Getzenberg RH, Wheeler MA, et al. A phase 2 cancer chemoprevention biomarker trial of isoflavone G-2535 (genistein) in presurgical bladder cancer patients. Cancer Prev Res (Phila). 2012; 5:621-630.
46. Wang YW, Chuang JJ, Chang TY, Won SJ, Tsai HW, Lee CT, Cheng HL, Tzai TS, Liu HS, Chow NH. Antiangiogenesis as the novel mechanism for justicidin A in the anticancer effect on human bladder cancer. Anticancer Drugs. 2015; 26:428-436.
47. Lee ST, Lu MH, Chien LH, Wu TF, Huang LC, Liao GI. Suppression of urinary bladder urothelial carcinoma cell by the ethanol extract of pomegranate fruit through cell cycle arrest and apoptosis. BMC Complement Altern Med. 2013; 13:364.
48. Kemberling JK, Hampton JA, Keck RW, Gomez MA, Selman SH. Inhibition of bladder tumor growth by the green tea derivative epigallocatechin-3-gallate. J Urol. 2003; 170:773-776.
49. Komori A, Yatsunami J, Okabe S, Abe S, Hara K, Suganuma M, Kim SJ, Fujiki H. Anticarcinogenic activity of green tea polyphenols. Jpn J Clin Oncol. 1993; 23:186-190.
50. Lu QY, Jin YS, Pantuck A, Zhang ZF, Heber D, Belldegrun A, Brooks M, Figlin R, Rao J. Green tea extract modulates actin remodeling via Rho activity in an in vitro multistep carcinogenic model. Clin Cancer Res. 2005; 11:1675-1683.
51. Philips BJ, Coyle CH, Morrisroe SN, Chancellor MB, Yoshimura N. Induction of apoptosis in human bladder cancer cells by green tea catechins. Biomed Res. 2009; 30:207-215.
52. Fimognari C, Hrelia P. Sulforaphane as a promising molecule for fighting cancer. Mutat Res. 2007; 635:90-104.
53. Shapiro TA, Fahey JW, Wade KL, Stephenson KK, Talalay P. Human metabolism and excretion of cancer chemoprotective glucosinolates and isothiocyanates of cruciferous vegetables. Cancer Epidemiol Biomarkers Prev. 1998; 7:1091-1100.
54. Zhang Y, Talalay P, Cho CG, Posner GH. A major inducer of anticarcinogenic protective enzymes from broccoli: isolation and elucidation of structure. Proc Natl Acad Sci U S A. 1992; 89:2399-2403.
55. Michaud DS, Spiegelman D, Clinton SK, Rimm EB, Willett WC, Giovannucci E. Prospective study of dietary supplements, macronutrients, micronutrients, and risk of bladder cancer in US men. Am J Epidemiol. 2000; 152:1145-1153.
56. Michaud DS, Clinton SK, Rimm EB, Willett WC, Giovannucci E. Risk of bladder cancer by geographic region in a U.S. cohort of male health professionals. Epidemiology. 2001; 12:719-726.
57. Michaud DS, Spiegelman D, Clinton SK, Rimm EB, Willett WC, Giovannucci EL. Fruit and vegetable intake and incidence of bladder cancer in a male prospective cohort. J Natl Cancer Inst. 1999; 91:605-613.
58. Liu B, Mao Q, Lin Y, Zhou F, Xie L. The association of cruciferous vegetables intake and risk of bladder cancer: a meta-analysis. World J Urol. 2013; 31:127-133.
59. Brüsewitz G, Cameron BD, Chasseaud LF, Gorler K, Hawkins DR, Koch H, Mennicke WH. The metabolism of benzyl isothiocyanate and its cysteine conjugate. Biochem J. 1977; 162:99-107.
60. Mennicke WH, Gorler K, Krumbiegel G, Lorenz D, Rittmann N. Studies on the metabolism and excretion of benzyl isothiocyanate in man. Xenobiotica. 1988; 18:441-447.
61. Ye L, Dinkova-Kostova AT, Wade KL, Zhang Y, Shapiro TA, Talalay P. Quantitative determination of dithiocarbamates in human plasma, serum, erythrocytes and urine: pharmacokinetics of broccoli sprout isothiocyanates in humans. Clin Chim Acta. 2002; 316:43-53.
62. Egner PA, Chen JG, Wang JB, Wu Y, Sun Y, Lu JH, Zhu J, Zhang YH, Chen YS, Friesen MD, Jacobson LP, Munoz A, Ng D, et al. Bioavailability of Sulforaphane from two broccoli sprout beverages: results of a short-term, cross-over clinical trial in Qidong, China. Cancer Prev Res (Phila). 2011; 4:384-395.
63. Kassahun K, Davis M, Hu P, Martin B, Baillie T. Biotransformation of the naturally occurring isothiocyanate sulforaphane in the rat: identification of phase I metabolites and glutathione conjugates. Chem Res Toxicol. 1997; 10:1228-1233.
64. Hienert G, Latal D, Rummelhardt S. Urological aspects of surgical management for metastatic renal cell cancer. Semin Surg Oncol. 1988; 4:137-138.
65. Zhang Y. Role of glutathione in the accumulation of anticarcinogenic isothiocyanates and their glutathione conjugates by murine hepatoma cells. Carcinogenesis. 2000; 21:1175-1182.
66. Zhang Y. Molecular mechanism of rapid cellular accumulation of anticarcinogenic isothiocyanates. Carcinogenesis. 2001; 22:425-431.
67. Zhang Y. The molecular basis that unifies the metabolism, cellular uptake and chemopreventive activities of dietary isothiocyanates. Carcinogenesis. 2012; 33:2-9.
68. Abbaoui B, Riedl KM, Ralston RA, Thomas-Ahner JM, Schwartz SJ, Clinton SK, Mortazavi A. Inhibition of bladder cancer by broccoli isothiocyanates sulforaphane and erucin: characterization, metabolism, and interconversion. Mol Nutr Food Res. 2012; 56:1675-1687.
69. Bricker GV, Riedl KM, Ralston RA, Tober KL, Oberyszyn TM, Schwartz SJ. Isothiocyanate metabolism, distribution, and interconversion in mice following consumption of thermally processed broccoli sprouts or purified sulforaphane. Mol Nutr Food Res. 2014; 58:1991-2000.
70. Munday R, Munday CM. Induction of phase II detoxification enzymes in rats by plant-derived isothiocyanates: comparison of allyl isothiocyanate with sulforaphane and related compounds. J Agric Food Chem. 2004; 52:1867-1871.
71. Munday R, Mhawech-Fauceglia P, Munday CM, Paonessa JD, Tang L, Munday JS, Lister C, Wilson P, Fahey JW, Davis W, Zhang Y. Inhibition of urinary bladder carcinogenesis by broccoli sprouts. Cancer Res. 2008; 68:1593-1600.
72. Bogaards JJ, Verhagen H, Willems MI, van Poppel G, van Bladeren PJ. Consumption of Brussels sprouts results in elevated alpha-class glutathione S-transferase levels in human blood plasma. Carcinogenesis. 1994; 15:1073-1075.
73. Cipolla BG, Mandron E, Lefort JM, Coadou Y, Della Negra E, Corbel L, Le Scodan R, Azzouzi AR, Mottet N. Effect of Sulforaphane in Men with Biochemical Recurrence after Radical Prostatectomy. Cancer Prev Res (Phila). 2015; 8:712-719.
74. Houghton CA, Fassett RG, Coombes JS. Sulforaphane: translational research from laboratory bench to clinic. Nutr Rev. 2013; 71:709-726.
75. Saha S, Hollands W, Teucher B, Needs PW, Narbad A, Ortori CA, Barrett DA, Rossiter JT, Mithen RF, Kroon PA. Isothiocyanate concentrations and interconversion of sulforaphane to erucin in human subjects after consumption of commercial frozen broccoli compared to fresh broccoli. Mol Nutr Food Res. 2012; 56:1906-1916.
76. Atwell LL, Hsu A, Wong CP, Stevens JF, Bella D, Yu TW, Pereira CB, Lohr CV, Christensen JM, Dashwood RH, Williams DE, Shannon J, Ho E. Absorption and chemopreventive targets of sulforaphane in humans following consumption of broccoli sprouts or a myrosinase-treated broccoli sprout extract. Mol Nutr Food Res. 2015; 59:424-433.
77. Veeranki OL, Bhattacharya A, Marshall JR, Zhang Y. Organ-specific exposure and response to sulforaphane, a key chemopreventive ingredient in broccoli: implications for cancer prevention. Br J Nutr. 2013; 109:25-32.
78. Jo GH, Kim GY, Kim WJ, Park KY, Choi YH. Sulforaphane induces apoptosis in T24 human urinary bladder cancer cells through a reactive oxygen species-mediated mitochondrial pathway: the involvement of endoplasmic reticulum stress and the Nrf2 signaling pathway. Int J Oncol. 2014; 45:1497-1506.
79. Nijhoff WA, Grubben MJ, Nagengast FM, Jansen JB, Verhagen H, van Poppel G, Peters WH. Effects of consumption of Brussels sprouts on intestinal and lymphocytic glutathione S-transferases in humans. Carcinogenesis. 1995; 16:2125-2128.
80. Fahey JW, Zhang Y, Talalay P. Broccoli sprouts: an exceptionally rich source of inducers of enzymes that protect against chemical carcinogens. Proc Natl Acad Sci U S A. 1997; 94:10367-10372.
81. Hwang ES, Jeffery EH. Effects of different processing methods on induction of quinone reductase by dietary broccoli in rats. J Med Food. 2004; 7:95-99.
82, Talalay P, Fahey JW. Phytochemicals from cruciferous plants protect against cancer by modulating carcinogen metabolism. J Nutr. 2001; 131:3027S-3033S.
83. James D, Devaraj S, Bellur P, Lakkanna S, Vicini J, Boddupalli S. Novel concepts of broccoli sulforaphanes and disease: induction of phase II antioxidant and detoxification enzymes by enhanced-glucoraphanin broccoli. Nutr Rev. 2012; 70:654-665.
84. Lee YM, Cho HJ, Ponnuraj SP, Kim J, Kim JS, Kim SG, Park JH. Phenethyl isothiocyanate inhibits 12-O-tetradecanoylphorbol-13-acetate-induced inflammatory responses in mouse skin. J Med Food. 2011; 14:377-385.
85. Shan Y, Wu K, Wang W, Wang S, Lin N, Zhao R, Cassidy A, Bao Y. Sulforaphane down-regulates COX-2 expression by activating p38 and inhibiting NF-kappaB-DNA-binding activity in human bladder T24 cells. Int J Oncol. 2009; 34:1129-1134.
86. Youn HS, Kim YS, Park ZY, Kim SY, Choi NY, Joung SM, Seo JA, Lim KM, Kwak MK, Hwang DH, Lee JY. Sulforaphane suppresses oligomerization of TLR4 in a thiol-dependent manner. J Immunol. 2010; 184:411-419.
87. Tang L, Zhang Y. Mitochondria are the primary target in isothiocyanate-induced apoptosis in human bladder cancer cells. Mol Cancer Ther. 2005; 4:1250-1259.
88. Park HS, Han MH, Kim GY, Moon SK, Kim WJ, Hwang HJ, Park KY, Choi YH. Sulforaphane induces reactive oxygen species-mediated mitotic arrest and subsequent apoptosis in human bladder cancer 5637 cells. Food Chem Toxicol. 2014; 64:157-165.
89. Singh SV, Singh K. Cancer chemoprevention with dietary isothiocyanates mature for clinical translational research. Carcinogenesis. 2012; 33:1833-1842.
90. Zhang Y. Allyl isothiocyanate as a cancer chemopreventive phytochemical. Mol Nutr Food Res. 2010; 54:127-135.
91. Choi S, Singh SV. Bax and Bak are required for apoptosis induction by sulforaphane, a cruciferous vegetable-derived cancer chemopreventive agent. Cancer Res. 2005; 65:2035-2043.
92. Choi S, Lew KL, Xiao H, Herman-Antosiewicz A, Xiao D, Brown CK, Singh SV. D,L-Sulforaphane-induced cell death in human prostate cancer cells is regulated by inhibitor of apoptosis family proteins and Apaf-1. Carcinogenesis. 2007; 28:151-162.
93. Gibbs A, Schwartzman J, Deng V, Alumkal J. Sulforaphane destabilizes the androgen receptor in prostate cancer cells by inactivating histone deacetylase 6. Proc Natl Acad Sci U S A. 2009; 106:16663-16668.
94. Gamet-Payrastre L, Li P, Lumeau S, Cassar G, Dupont MA, Chevolleau S, Gasc N, Tulliez J, Terce F. Sulforaphane, a naturally occurring isothiocyanate, induces cell cycle arrest and apoptosis in HT29 human colon cancer cells. Cancer Res. 2000; 60:1426-1433.
95. Suppipat K, Park CS, Shen Y, Zhu X, Lacorazza HD. Sulforaphane induces cell cycle arrest and apoptosis in acute lymphoblastic leukemia cells. PLoS One. 2012; 7:e51251.
96. Shan Y, Sun C, Zhao X, Wu K, Cassidy A, Bao Y. Effect of sulforaphane on cell growth, G(0)/G(1) phase cell progression and apoptosis in human bladder cancer T24 cells. Int J Oncol. 2006; 29:883-888.
97. Tang L, Zhang Y. Dietary isothiocyanates inhibit the growth of human bladder carcinoma cells. J Nutr. 2004; 134:2004-2010.
98. Zhang Y, Tang L, Gonzalez V. Selected isothiocyanates rapidly induce growth inhibition of cancer cells. Mol Cancer Ther. 2003; 2:1045-1052.
99. Dang YM, Huang G, Chen YR, Dang ZF, Chen C, Liu FL, Guo YF, Xie XD. Sulforaphane inhibits the proliferation of the BIU87 bladder cancer cell line via IGFBP-3 elevation. Asian Pac J Cancer Prev. 2014; 15:1517-1520.
100. Dickinson SE, Melton TF, Olson ER, Zhang J, Saboda K, Bowden GT. Inhibition of activator protein-1 by sulforaphane involves interaction with cysteine in the cFos DNA-binding domain: implications for chemoprevention of UVB-induced skin cancer. Cancer Res. 2009; 69:7103-7110.
101. Forster T, Rausch V, Zhang Y, Isayev O, Heilmann K, Schoensiegel F, Liu L, Nessling M, Richter K, Labsch S, Nwaeburu CC, Mattern J, Gladkich J, et al. Sulforaphane counteracts aggressiveness of pancreatic cancer driven by dysregulated Cx43-mediated gap junctional intercellular communication. Oncotarget. 2014; 5:1621-1634. doi: 10.18632/oncotarget.1764.
102. Clarke JD, Hsu A, Riedl K, Bella D, Schwartz SJ, Stevens JF, Ho E. Bioavailability and inter-conversion of sulforaphane and erucin in human subjects consuming broccoli sprouts or broccoli supplement in a cross-over study design. Pharmacol Res. 2011; 64:456-463.
103. Wang F, Shan Y. Sulforaphane retards the growth of UM-UC-3 xenographs, induces apoptosis, and reduces survivin in athymic mice. Nutr Res. 2012; 32:374-380.
104. Akagi K, Sano M, Ogawa K, Hirose M, Goshima H, Shirai T. Involvement of toxicity as an early event in urinary bladder carcinogenesis induced by phenethyl isothiocyanate, benzyl isothiocyanate, and analogues in F344 rats. Toxicol Pathol. 2003; 31:388-396.
105. Alumkal JJ, Slottke R, Schwartzman J, Cherala G, Munar M, Graff JN, Beer TM, Ryan CW, Koop DR, Gibbs A, Gao L, Flamiatos JF, Tucker E, et al. A phase II study of sulforaphane-rich broccoli sprout extracts in men with recurrent prostate cancer. Invest New Drugs. 2015; 33:480-489.
106. Block KI, Gyllenhaal C, Lowe L, Amedei A, Amin AR, Amin A, Aquilano K, Arbiser J, Arreola A, Arzumanyan A, Ashraf SS, Azmi AS, Benencia F, et al. Designing a broad-spectrum integrative approach for cancer prevention and treatment. Semin Cancer Biol. 2015; 35 Suppl:S276-304.
107. Knowles MA, Hurst CD. Molecular biology of bladder cancer: new insights into pathogenesis and clinical diversity. Nat Rev Cancer. 2015; 15:25-41.
108. Tong YH, Zhang B, Fan YM, Lin NM. Keap1-Nrf2 pathway: A promising target towards lung cancer prevention and therapeutics. Chronic Diseases and Translational Medicine. 2015; 1:175-186.
109. Sharma R, Sharma A, Chaudhary P, Sahu M, Jaiswal S, Awasthi S, Awasthi YC. Role of 4-hydroxynonenal in chemopreventive activities of sulforaphane. Free Radic Biol Med. 2012; 52:2177-2185.
110. Houghton CA, Fassett RG, Coombes JS. Sulforaphane and Other Nutrigenomic Nrf2 Activators: Can the Clinician’s Expectation Be Matched by the Reality? Oxid Med Cell Longev. 2016; 2016:7857186.
111. Lan A, Li W, Liu Y, Xiong Z, Zhang X, Zhou S, Palko O, Chen H, Kapita M, Prigge JR, Schmidt EE, Sun Z, Chen X, et al. Chemoprevention of oxidative stress-associated oral carcinogenesis by sulforaphane depends on NRF2 and the isothiocyanate moiety. Oncotarget. 2016; 7:53502-53514. doi: 10.18632/oncotarget.10609.
112. Sestili P, Fimognari C. Cytotoxic and Antitumor Activity of Sulforaphane: The Role of Reactive Oxygen Species. Biomed Res Int. 2015; 2015:402386.
113. Juengel E, Maxeiner S, Rutz J, Justin S, Roos F, Khoder W, Tsaur I, Nelson K, Bechstein WO, Haferkamp A, Blaheta RA. Sulforaphane inhibits proliferation and invasive activity of everolimus-resistant kidney cancer cells in vitro. Oncotarget. 2016; 7:85208-85219. doi: 10.18632/oncotarget.13421.
114. Rizzo VL, Levine CB, Wakshlag JJ. The effects of sulforaphane on canine osteosarcoma proliferation and invasion. Vet Comp Oncol. 2016.
115. Meeran SM, Patel SN, Tollefsbol TO. Sulforaphane causes epigenetic repression of hTERT expression in human breast cancer cell lines. PLoS One. 2010; 5:e11457.
116. Cheng YM, Tsai CC, Hsu YC. Sulforaphane, a Dietary Isothiocyanate, Induces G(2)/M Arrest in Cervical Cancer Cells through CyclinB1 Downregulation and GADD45beta/CDC2 Association. Int J Mol Sci. 2016; 17.
117. Islam SS, Mokhtari RB, Akbari P, Hatina J, Yeger H, Farhat WA. Simultaneous Targeting of Bladder Tumor Growth, Survival, and Epithelial-to-Mesenchymal Transition with a Novel Therapeutic Combination of Acetazolamide (AZ) and Sulforaphane (SFN). Target Oncol. 2016; 11:209-227.
118. Hudecova S, Markova J, Simko V, Csaderova L, Stracina T, Sirova M, Fojtu M, Svastova E, Gronesova P, Gronesova P, Pastorek M, Novakova M, Cholujova D, et al. Sulforaphane-induced apoptosis involves the type 1 IP3 receptor. Oncotarget. 2016; 7:61403-61418. doi: 10.18632/oncotarget.8968.
119. Yang L, Palliyaguru DL, Kensler TW. Frugal chemoprevention: targeting Nrf2 with foods rich in sulforaphane. Semin Oncol. 2016; 43:146-153.
120. Atwell LL, Zhang Z, Mori M, Farris PE, Vetto JT, Naik AM, Oh KY, Thuillier P, Ho E, Shannon J. Sulforaphane Bioavailability and Chemopreventive Activity in Women Scheduled for Breast Biopsy. Cancer Prev Res (Phila). 2015; 8:1184-1191.
121. Li QQ, Xie YK, Wu Y, Li LL, Liu Y, Miao XB, Liu QZ, Yao KT, Xiao GH. Sulforaphane inhibits cancer stem-like cell properties and cisplatin resistance through miR-214-mediated downregulation of c-MYC in non-small cell lung cancer. Oncotarget. 2017; 8:12067-12080. doi: 10.18632/oncotarget.14512.
122. Taguchi K, Motohashi H, Yamamoto M. Molecular mechanisms of the Keap1-Nrf2 pathway in stress response and cancer evolution. Genes Cells. 2011; 16:123-140.
123. Traka M, Gasper AV, Melchini A, Bacon JR, Needs PW, Frost V, Chantry A, Jones AM, Ortori CA, Barrett DA, Ball RY, Mills RD, Mithen RF. Broccoli consumption interacts with GSTM1 to perturb oncogenic signaling pathways in the prostate. PLoS One. 2008; 3:e2568.
124. Wang L, Tian Z, Yang Q, Li H, Guan H, Shi B, Hou P, Ji M. Sulforaphane inhibits thyroid cancer cell growth and invasiveness through the reactive oxygen species-dependent pathway. Oncotarget. 2015; 6:25917-25931. doi: 10.18632/oncotarget.4542.
125. Fahey JW, Wade KL, Wehage SL, Holtzclaw WD, Liu H, Talalay P, Fuchs E, Stephenson KK. Stabilized sulforaphane for clinical use: Phytochemical delivery efficiency. Mol Nutr Food Res. 2017; 61.